Influence of Solar Radiation on the Morphometric and Biochemical Traits of Rocket (Eruca sativa Mill.) and Sicilian Black Sprouting Broccoli (Brassica oleracea var. italica Plenck) Microgreens
Received Date: January 07, 2024 Accepted Date: February 07, 2024 Published Date: February 10, 2024
doi: 10.17303/jacs.2024.3.101
Citation: Donata Arena, Hajer Ben Ammar, Nikola Major, Tvrtko Karlo Kovacevic, Smiljana Goreta Banet al. (2024) Influence of Solar Radiation on the Morphometric and Biochemical Traits of Rocket (Eruca sativa Mill.) and Sicilian Black Sprouting Broccoli (Brassica oleracea var. italica Plenck) Microgreens. J Adv Agron Crop Sci 3: 1-17
Abstract
The impact of light on plant growth and biochemical profile is a fundamental aspect of plant physiology. This study aimed to investigate how solar radiation affects the growth and biochemical composition of microgreens, including two commercial rocket genotypes (Eruca sativa Mill. var. darkita and Eruca sativa Mill. var. lobata, RD1 and RD2, respectively) and a Sicilian Black sprouting broccoli (Brassica oleracea var. italica Plenck, Broccolo nero, BR), under varying levels of solar radiation (control (SR100) and stress (SR40)) using black shade nets. The plantlets were analyzed for their main morphometric traits such as weight, hypocotyl length, cotyledon and leaf dimensions, and biochemical parameters including total flavonoid content (TFC), DPPH (2,2-diphenyl-1-picrylhydrazyl) scavenging activity, and FRAP (ferric reducing antioxidant power). Additionally, we determined the total glucosinolates content, sucrose, fructose, and fructooligosaccharides (FOS). BR exhibited the highest antioxidant capacity compared to the other genotypes tested. Regarding the sugar profile, significant variations were observed, with RD1 and RD2 exhibiting the highest amount of sugar. Reduced solar radiation levels led to a decrease in fructooligosaccharides (FOS) levels, while sucrose, glucose, and fructose increased. TFC was particularly influenced by control conditions. This study enhances our understanding of microgreens, providing insights into their response to solar radiation variations and highlighting their potential commercial use in the market.
Keywords: Brassicaceae; Plant Growth Stage; Shade Net; Antioxidant Activity; Sugars
Introduction
Over the past three decades, consumer preferences have undergone a notable shift towards natural foods, particularly those rich in bioactive compounds. This trend reflects a growing interest in maintaining a balanced and healthy diet [1]. This is achieved through innovation in the development of new products, with a focus on adding value to agro-biodiversity at the local level. By doing so, there is a dual benefit of reducing environmental impacts while concurrently offering ready-to-eat options for reformulated vegetables [2]. The food supply chains have responded to this shift in consumer behavior by introducing new, competitive, and innovative products. Among these offerings are scientifically reformulated items, such as edible sprouts, microgreens, and baby leaf salads [3]. Microgreens are young edible plants harvested shortly after germination, typically within a few weeks. They fall between the size of sprouts and baby greens, typically reaching heights of 1-3 inches. Cultivated from a diverse range of vegetables such as kale, spinach, radish, broccoli, and more, microgreens are harvested at the stage when their first true leaves emerge, hence the term "microgreens" [4].
The Brassicaceae family encompasses a diverse range of cruciferous vegetables, which includes well-known varieties like broccoli, kale, cauliflower, cabbage, kohlrabi, and various wild rocket species valued for their edible leaves, stems, or flower heads [5]. This extensive diversity can be traced back to an ancient domestication process that occurred in the Mediterranean basin and involved the cultivation of crop wild relatives (CWRs) [6,7]. Brassica oleracea L. stands out as a model organism in plant science research, maintaining consistent scientific interest due to its contributions to essential genetic [8,9] and physiological processes [10]. Notably, Brassica oleracea L. possesses a distinctive biochemical profile abundant in bioactive compounds with antioxidant properties, including glucosinolates, polyphenols, ascorbic acid, and vitamins [11,12]. Similarly, Eruca sativa, commonly known as rocket salad, has been a focal point in numerous studies, showcasing its high content of bioactive compounds such as glucosinolates[13], carotenoids, vitamin C, fibers, and flavonoids [14,15].
The agronomic characteristics and the qualitative and quantitative aspects of bioactive compounds are subject to modulation by different factors, including plant organ specificity, developmental stages, and external influencers such as environmental stresses and biological stimuli [16-18]. Predominant among the abiotic stressors are alterations in light irradiance, temperature fluctuations, intermittent periods of aridity, and heightened salinity levels. In response to adverse environmental conditions, Brassica crops adeptly increase their antioxidant defense mechanisms, orchestrating a concerted response to alleviate the deleterious effects of oxidative stress [19,20]. This intricate interplay of environmental factors and biochemical responses underscores the dynamic relationship between plant physiology and external stressors, particularly in the context of Brassica crops and their resilience against oxidative challenges [21].
Light stands as a pivotal environmental determinant with a substantial impact on the morphological and biochemical attributes of broccoli (Brassica oleracea var. italica) [22]. Broccoli plants intricately adapt to fluctuating solar radiation conditions through the process of photomorphogenesis [23]. Biochemically, solar radiation profoundly influences the accumulation of pigments and bioactive compounds in Brassica crops. Optimal light levels foster an increase in chlorophyll content, facilitating efficient photosynthesis. Solar radiation serves as a catalyst for the biosynthesis of carotenoids, including beta-carotene, lutein, zeaxanthin, and anthocyanins [24].
The investigation entails a meticulous analysis of morphometric characteristics and biochemical profiles across distinct genotypes of commercially relevant rocket plants, namely Eruca sativa Mill. var. darkita and Eruca sativa Mill. var. lobata, alongside Sicilian Black sprouting broccoli (Brassica oleracea var. italica Plenck, Broccolo nero), at various growth stages encompassing sprouts, microgreens, and baby leaves. Evaluation parameters include principal morphometric factors, as well as the biochemical fingerprint in correlation with the considered experimental variables. Key biochemical parameters such as total phenolic content (TPC), total flavonoid content (TFC), and various antioxidant assays, including DPPH (2,2-diphenyl-1-picrylhydrazyl), ORAC (oxygen radical absorbance capacity), and FRAP (ferric reducing antioxidant power), were meticulously determined to comprehensively elucidate the intricate interplay between solar radiation and the biochemical composition of these plants.
Material and Methods
Experimental Design
The experiment compared two solar radiation conditions: SR100 for 100% solar radiation (control) and SR40 for 40% solar radiation (stress). It employed a split-plot randomized design with two experimental factors: different solar radiation levels (control and stress) and genotype (GE). Each experimental treatment was replicated three times. The experimental trial took place in a cold greenhouse on September 11, 2023, in Catania, Italy (37°31′10″ N 15°04′18″ E; 105 m above sea level (m a.s.l.)), following organic practices. The seeds of the Sicilian landrace Broccolo nero (BR, Brassica oleracea var. italica Plenck) were sourced from the Di3A active brassica genebank collection, while the seeds of two rocket genotypes (RD1 and RD2, Eruca sativa Mill. var. darkita and Eruca sativa Mill. var. lobata, respectively) were provided by S.A.I.S. S.p.A. seed company (Cesena, Italy). The seeds were sown in cellular trays filled with organic substrate Brill®semina bio (Geotec, Italy), and plantlets were irrigated using standard techniques. A black polyethylene shade net (Arrigoni, Agrotextiles & Techtextiles): 3070NE SCIROCCO 75 (hole dimension 0.35x1.39 mm) was utilized, with the control plot left without a net. Light intensity was measured using a LI-COR Quantum/Radiometer/Photometer (Model LI-250 Light Meter). The plantlets were collected with the first appearance of the true leaf after 15 days from the sowing.
Morphometric Measurements
The plants characterized for their main morphometric traits: the weight of 10 individuals (W), hypocotyllength (HL), cotyledon length and width (CL and CW, respectively), for sprouts; in addition, number (N), length and width of the true leaves (LL and LW, respectively) for microgreens. measurements were performed using Epson Perfection V850 Pro (WinFolia Regular 2020). The plants, after the characterization, were frozen at -80°C, freeze-dried, and grounded to obtain a fine powder, and then utilized for biochemical analysis.
Phytochemical Measurements
The phytochemical extracts were prepared by homogenizing 50 mg of freeze-dried material with 1 mL of aqueous methanol (80/20, v/v) using 2.4 mm metal beads (Omni kit 19-670, Kennesaw, GA, USA) at room temperature for 1 minute at 5 m/s (OmniBeadRuptorElite, Kennesaw, GA, USA). The mixture was then centrifuged at 16,000 x g for 10 minutes (Centric 350, Tehtnica), and the supernatants were transferred to clean tubes and stored at -80°C until further analysis.
Total Antioxidant Capacity Measurement
Ferric Reducing Antioxidant Power (FRAP)
The FRAP assay involved mixing 100 µL of sample extract with 200 µL of FRAP reagent following Benzie et al.'s method [25]. After a 10-minute incubation at room temperature, absorbance was measured at 593 nm using a Tecan Infinite 200 Pro M Nano+ spectrophotometer (Männedorf, Switzerland). FRAP values were determined based on a Trolox+ calibration curve (y = 10.0044x − 0.094) constructed with serial Trolox dilutions (20, 40, 60, 80, 100 µM) exhibiting a high coefficient of determination (R² = 0.9995), expressed as µmol TE/g d.w.
1,1-diphenyl-2-picrylhydrazil scavenging (DPPH)
100 µL of sample extract was mixed with 200 µL of 0.1 mM DPPH reagent, following Brand-Williams et al.'s method [26]. After a 30-minute incubation at room temperature, absorbance was measured at 517 nm. Results were expressed as µmol TE/g d.w. using a Trolox standard curve (y = −15.147x + 15.186) derived from Trolox dilutions (20, 40, 60, 80, 100 µM) with a remarkable coefficient of determination (R² = 0.9999).
Total Flavonoid Content (TFC)
TFC was determined using the aluminum chloride colorimetric assay method described by Sembiring et al. [27] with adaptations. Absorbance was recorded at 415 nm after a 40-minute incubation period, and results were expressed as milligrams of Quercetin (QE)/g d.w. using a Quercetin standard curve (y= 7.1021x – 0.005) based on serial Quercetin dilutions (20 to 100 mg/L) with a high coefficient of determination (R²= 0.9999).
Total Glucosinolates Content (SEQ)
100 µL of the plant extract was combined with 100µL of a 0.2 M Na2PdCl4 for the determination of total glucosinolates content. After one minute, 100 µL of a 6% (w/v) sodium carbonate (Na2CO3) solution were added. The mixture then was incubated at 25 °C for 60 min, following absorbance measurement at 750 nm using a Tecan Infinite 200 Pro M Nano+ spectrophotometer (Männedorf, Switzerland). The quantification was carried out using a standard curve constructed with sinigrin (y = 3.7585x − 0.0035), involving serial dilutions ranging from 20 to 100 µg/mL. This calibration exhibited a high coefficient of determination (R²= 0.9997). The results were expressed in terms of gram of Sinigrin Equivalents (SEQ) per gram of dry weight (d.w.).
Soluble Sugar Analysis
150 mg of freeze-dried samples were homogenized with 2.4 mm metal beads for 1 min at 5 m/s in 3 mL of 80% methanol in water using a bead mill (Omni kit 19-670, Kennesaw, GA, USA) for 1 min at 5 m s-1 in 3 mL of 80% methanol in water using a bead mill (Omni Bead Ruptor Elite, Kennesaw, GA, USA). The samples were subsequently centrifuged for 5 min at 5000×g. The extracts were filtered through a 0.22 µm nylon filter. The analysis of sucrose, glucose, fructose, and inulin (FOS) content was carried out using an HPLC system with a system controller (Shimadzu CBM-40, Kyoto, Japan), a degassing unit (Shimadzu DGU-405, Kyoto, Japan), a solvent delivery unit (Shimadzu LC-20Ai, Kyoto, Japan), an autosampler (Shimadzu SIL-20AC, Kyoto, Japan), column oven (Shimadzu CTO-40S, Kyoto, Japan) and a refractive index detector (Shimadzu RID-20A, Kyoto, Japan). The chromatographic separation was carried out by injecting 10 µL of the sample on a 300 × 8 mm, 9 µm particle size, calcium cation exchange column (Ammerbuch, Germany) held at 80 °C using deionized water as the mobile phase (0.6 mL min-1, isocratic elution). The results were elaborated by comparing the retention times and peak areas of the investigated sugars to analytical standards for identification and quantification, respectively. The calibration curves were obtained with serial dilutions of inulin (0.05, 0.1, 0.5, 1.0, 5.0, 10.0 and 20.0 g L-1) (y = 114.85x - 1.743, coefficient of determination, R2 = 0.99999), sucrose (y = 130.79x − 0.3583, coefficient of determination,R2 = 0.99999), glucose (y = 132.36x − 2.101, coefficient of determination, R2= 0.9999). The results were given in gram/100 grams of dry weight (d.w.).
Statistical Analysis
The data are presented as mean ± standard deviation (SD) of replicates for each analysis. The statistical significance of morphometric trait differences was assessed using a two-way analysis of variance (ANOVA) in CoStat version 6.451 (CoHort Software, Birmingham, England). For biochemical profiles, a two-way ANOVA considering genotypes and stress conditions was conducted, followed by Tukey’s multiple comparisons test, performed in GraphPad Prism version 8.0 (GraphPad Software, Inc., San Diego, CA, USA). Statistical significance was defined at p-values < 0.05. Principal component analysis (PCA) was employed for multivariate analysis to elucidate and summarize sample differences using XLSTAT2018 software (Addinsoft, Paris, France).
Results
Climatic Conditions
Variations of the climatic conditions were observed among the different experimental thesis compared. The main climatic conditions were registered from the beginning of the trial (September 11, 2023), after 7 days from sowing (September 18, 2023) and at the end of the experiment (September 25, 2023).
The solar radiation intensity varied from 126.38 ±15.47 µmol s-1 m-2 for the control to 58.74 ± 0.62 µmol s-1 m-2for the stress (Figure 1).
Morphometric Parameters
For microgreens, no significant differences were observed for the weight. We ascertained a significant interaction between SR and GE for hypocotyl length, and its values varied from 5.08 ± 0.54 mm to 1.99 ± 0.15 mm, for BR SR40 and RD2 SR100, respectively (Table 1). The cotyledon length was significantly affected by the genotype, with values ranged from 9.10 to 10.45 mm, for RD1 and BR, respectively. A significant interaction was observed for cotyledon width, which value varied from 1.01 to 0.89 mm, for BR SR40 and SR100, respectively. No significant variations were ascertained for leaf length. The highest value was ascertained for RD1 SR100 (3.35 mm), whereas the lowest one was determined for BR SR40 (3.46 mm). For leaf width, we observed a significant interaction between SR and GE and its value varied from 0.22 to 0.38 for RD2 SR100 and BR SR100, respectively.
Phytochemical Measurements
The results of the FRAP activity analysis revealed a significant variation among the tested genotypes, as depicted in Figure 2. BR exhibited the highest antioxidant activity, with a value of 38.07 µmol TE/g d.w. under control conditions, which closely resembled the stress plot. Conversely, RD2 demonstrated an increase in FRAP activity under stress conditions, registering a value of 25.90 µmol TE/g d.w. compared to the control (Figure 2).
Regarding the DPPH activity, BR demonstrated the highest value under control conditions (19.44 µmol TE/g d.w.), more than double compared to RD1 and RD2, which recorded 10.55 and 6.85 µmol TE/g d.w. under control conditions, respectively. Similarly, RD2 exhibited an increase in DPPH activity under stress conditions (8.87 µmol TE/g d.w.) (Figure 3).
Regarding the DPPH activity, BR demonstrated the highest value under control conditions (19.44 µmol TE/g d.w.), more than double compared to RD1 and RD2, which recorded 10.55 and 6.85 µmol TE/g d.w. under control conditions, respectively. Similarly, RD2 exhibited an increase in DPPH activity under stress conditions (8.87 µmol TE/g d.w.) (Figure 3).
The total glucosinolate content exhibits a significant variation between genotype and stress condition, with values ranging from 8.28 to 4.95 g SEQ/g d.w. for BR and RD2, respectively, under control conditions (Figure 5). Remarkably, RD1 demonstrated a comparable value to RD2, while for RD2, the total glucosinolates content tended to increase under stress conditions, ranging from 4.95 to 5.73 g SEQ/g d.w. for control and stress conditions, respectively.
The total sugars content (g/1000 g d.w.) exhibited significant variations among the genotypes. RD1 showed the highest amount under stress conditions, with values ranging from 78.67 to 193.03 g/1000 g d.w. from SR40 to SR100, respectively (Figure 6). In contrast, for the other genotypes tested, we observed no significant variations between stress and control conditions.
The glucose content (g/1000 g d.w.) exhibited significant variations in relation to the experimental factors considered. The smallest reduction was observed for BR, with a value of 4.79 g/1000 g d.w. under stress conditions compared to the control. The glucose amount generally increased from the control to stress conditions for the other genotypes tested, with values ranging from 26.26 to 38.22 g/1000 g d.w. for RD1 and RD2, respectively (Figure 7).
The FOS amount was significantly influenced by the experimental factors genotype and light effect (Figure 8). RD1 exhibited the lowest content under control conditions (50.13 g/1000 g d.w.) compared to the stress condition. RD2 and BR showed similar FOS amounts across both conditions studied.
The sucrose amount varied significantly in relation to solar radiation, as previously mentioned for glucose. The sucrose content ranged from 2.85 to 16.26 g/1000g d.w. in relation to solar radiation, with SR100 RD2 and SR40 BR exhibiting these values, respectively (Figure 9). Generally, the sucrose amount increased from stress to control conditions for all genotypes tested.
The correlation matrix provides a comprehensive overview of the associations between antioxidant activities (DPPH, FRAP), total flavonoid content (TFC), total glucosinolates (TGLSs), and sugar profiles (glucose, fructose, sucrose, and FOS) considering the microgreens stage. The heatmaps in Figure 10 depict the Pearson correlation between the different biochemical compounds. Regarding the control condition, DPPH demonstrates a strong positive correlation with FRAP (r=0.96), TGLSs (r=1.00). Additionally, DPPH exhibits notably strong positive correlations with FOS (r=0.47) and total sugars (r=0.32). TFC shows a strong positive correlation with sucrose (r=0.94) and glucose (r=0.86). Furthermore, FOS displays a positive correlation with the antioxidant assays, while it is negatively correlated with sucrose (r= -0.14) and glucose (r= -0.33). For the stress plot, DPPH and FRAP exhibit a strong positive correlation (r=0.98) (Figure 11). Additionally, DPPH shows a strong positive correlation with TGLSs and FOS (r=1.00). Fructo-oligosaccharides (FOS) share a negative correlation with TFC (r= -0.53), sucrose (r= -0.47), and glucose (r=-0.79). Total sugars and FOS are positively correlated (r=0.31) (Figure 11).
The distribution of the studied accessions along the PC1 and PC2 axes reveals that the first axis accounts for 61.11% of the total variation, while the second axis explains 30.96% of the total variability (Figure 12). PC1 is positively associated with antioxidant parameters such as DPPH, FRAP, total glucosinolates (TGLSs), FOS, and total sugars, indicating a higher antioxidant activity. Conversely, PC2 is positively correlated with total flavonoid content (TFC), sucrose, and glucose. Under both stress and control conditions, BR demonstrates a significant positive correlation with PC1, indicating a higher antioxidant activity compared to the sugars profile. BR consistently exhibits a preference for antioxidants across different solar radiation levels at the microgreens stage. In contrast, RD2 in both experimental conditions shows a correlation with TFC, particularly with sugars such as sucrose and glucose, suggesting a preference for characteristics linked to carbohydrates over antioxidant activities (Figure 12). RD1 displays a different trend: under stress conditions, it shows an association with TFC, sucrose, and glucose similar to RD2. However, under control conditions, its profile differs entirely. The distribution of accessions along the principal component axes provides valuable insights into the relationship between biochemical properties and plant responses to solar radiation. The different associations observed between genotypes and biochemical traits underscore the genotype-specific regulation of these compounds, contributing to our understanding of the biochemical diversity within cultivars.
Discussion
Different species and cultivars have been evaluated as potential candidates for space food production, with a focus on factors like nutritional value, resource efficiency, and adaptability to controlled environments [28]. Therefore, understanding how environmental variables interact to influence the growth, development, and phytochemical production of microgreens becomes crucial. The findings of this study provide valuable insights into the morphometric parameters and phytochemical composition of microgreens under solar radiation stress conditions, highlighting genotype-specific responses.
The morphometric analysis revealed significant variations in hypocotyl length, cotyledon length and width, and leaf width among the different genotypes concerning the stress conditions. Interestingly, the interaction between solar radiation and genotype influenced hypocotyl and cotyledon dimensions. This suggests that environmental factors, such as solar radiation, interact with genetic traits to modulate morphological characteristics in microgreens. Understanding these interactions is crucial for optimizing cultivation practices and enhancing crop yield and quality [29]. The observed variations in leaf width indicate genotype-specific responses to stress, with certain genotypes exhibiting wider leaves under stress conditions. This could be attributed to genetic factors influencing leaf development and adaptation to environmental stressors [30].
Phytochemical analysis revealed significant genotype-specific variations in antioxidant activities, total flavonoid content (TFC), total glucosinolate content, and sugar profiles under stress conditions. Notably, some genotypes exhibited increased antioxidant activity and phytochemical content in response to stress, suggesting a potential adaptive mechanism to enhance stress resilience and nutritional quality. The observed variations in antioxidant activities, particularly FRAP and DPPH, reflect the genotypic diversity in microgreens and their ability to scavenge free radicals under stress conditions. These findings underscore the crucial role of genotype selection in breeding programs targeted at enhancing the antioxidant capacity and nutritional value of microgreens. Duan et al. emphasize the pivotal significance of genetic variation in this regard [31]. Modulating light intensity can positively impact the nutraceutical values of plants [32]. Even under high light intensity, which may be perceived as stress by plants, they can synthesize antioxidant compounds to mitigate the formation of reactive oxygen species (ROS) [33].
The increase in TFC and total glucosinolate content under stress conditions suggests a dynamic response to environmental cues, potentially mediated by stress-responsive signaling pathways, our findings align with prior research, as studies by Huseby et al. [34] and Perez-Balibrea et al. [35].have also reported a significant reduction in the concentration of glucosinolates in plants grown in darkness compared to those grown in light conditions. Understanding the genetic and biochemical basis of these responses could facilitate the development of stress-tolerant varieties with enhanced phytochemical profiles.
The variations in sugar profiles, including glucose, fructose, sucrose, and fructo-oligosaccharides (FOS), highlight genotype-specific metabolic responses to stress. The observed increase in glucose content in certain genotypes under stress conditions may serve as a carbon source for energy metabolism and stress adaptation as reported by Kuai et al [36]. Similarly, the differential accumulation of sucrose and FOS underscores the complexity of carbohydrate metabolism in microgreens under stress. The findings of this study have implications for breeding programs and cultivation practices aimed at improving the nutritional quality and stress resilience of microgreens. Understanding the genetic basis of morphometric and phytochemical responses to stress can inform targeted breeding strategies to develop stress-tolerant varieties with enhanced nutritional profiles [37].
In controlled conditions, significant correlations were observed, notably between DPPH and FRAP (r=0.96), as well as with total GLSs (r=1.00), indicating a synchronized response in antioxidant defence mechanisms. These findings align with the research conducted by Major et al [38]. Equally intriguing were the positive correlations of DPPH with FOS (r=0.47) and total sugars (r=0.32), indicating potential crosstalk between antioxidant pathways and sugar metabolism. The strong positive correlations of TFC with sucrose (r=0.94) and glucose (r=0.86) underscore the role of flavonoids in carbohydrate regulation and energy metabolism. Conversely, the negative correlations of FOS with sucrose and glucose hint at a regulatory role of fructooligosaccharides in sugar homeostasis. Transitioning to stress conditions, the robust positive correlation between DPPH and FRAP (r=0.98) persisted, indicating a conserved antioxidative response even under adverse environmental conditions. Additionally, DPPH exhibited significant positive associations with TGLSs and FOS (r=1.00), implying a potential link between antioxidant capacity and glucosinolate production. Notably, FOS displayed negative correlations with TFC (r= -0.53), sucrose (r= -0.47), and glucose (r= -0.79), suggesting a trade-off between fructooligosaccharide accumulation and other biochemical pathways under stress. However, the positive correlation between total sugars and FOS (r=0.31) hints at a coordinated stress response, possibly through osmotic regulation or signaling mechanisms. Comparing the effects of light stress on the biochemical parameters elucidates distinctive responses within microgreens. Under control conditions, where light stress is absent, microgreens exhibit robust antioxidant defense mechanisms, as evidenced by strong positive correlations between DPPH and FRAP, indicating a coordinated response to neutralize reactive oxygen species. Moreover, the positive correlations of DPPH with TGLSs and FOS suggest a potential enhancement of antioxidant capacity and secondary metabolite production in the absence of stress. Conversely, under light stress conditions, microgreens appear to prioritize antioxidant defense mechanisms, as indicated by the high positive correlation between DPPH and FRAP. This suggests an adaptive response to mitigate oxidative damage caused by increased light intensity. Additionally, the significant positive associations between DPPH and TGLSs, as well as FOS, underscore the integration of antioxidant pathways with stress-responsive metabolic processes, potentially enhancing the plants' resilience to environmental challenges. Interestingly, the negative correlations observed between FOS and TFC, sucrose, and glucose under light stress suggest a trade-off between fructooligosaccharide accumulation and other biochemical pathways involved in growth and development. This implies that microgreens may allocate resources away from sugar metabolism towards stressresponsive pathways, such as antioxidant defense and secondary metabolite biosynthesis, under light stress conditions. Future research will focus on elucidating the molecular mechanisms underlying genotype-specific responses to stress in microgreens. Transcriptomic and metabolomic analyses could provide insights into the genetic pathways and metabolic networks regulating morphological and phytochemical traits under stress conditions.
Conclusion
In conclusion, our study provides valuable insights into the mechanisms governing plant responses to solar radiation, a critical factor influencing crop yield and sustainability. The observed variations in hypocotyl length, cotyledon dimensions, and leaf morphology serve as important indicators of the impact of solar radiation on plant development. Moreover, our investigation into phenolic content and antioxidant capacities, including Fructooligosaccharides (FOS), reveals genotype-specific regulation of these compounds, thereby enriching our understanding of biochemical diversity within cultivars. These findings have significant implications for the development of strategies aimed at enhancing crop productivity amidst evolving environmental dynamics. By elucidating the relationship between genetic and environmental interactions, we aim to contribute to the advancement of agricultural sustainability and productivity in microgreens, ultimately fostering the development of a more resilient and efficient agricultural landscape for future generations.
Author Contributions
Conceptualization, F.B., D.A., L.S.R. and G.B.S.; methodology, F.B., G.B.S., M.N. and L.S.R.; software, D.A. and H.B.A.; validation, F.B., L.S.R. and G.B.S.; formal analysis, F.B., D.A., H.B.A., K.K.T.; investigation, D.A., M.N., T.S., and K.K.T.; resources, F.B.; data curation, F.B., L.S.R., G.B.S., H.B.A and D.A.; writing—original draft preparation, D.A. and H.B.A.; writing—review and editing, F.B., G.B.S. and L.S.R.; visualization, F.B., G.B.S. and L.S.R.; supervision, F.B. and G.B.S.; project administration, F.B.; funding acquisition, F.B. All authors have read and agreed to the published version of the manuscript.
Funding Sources
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
- Cena H, Calder PC (2020) Defining a healthy diet: evidence for the role of contemporary dietary patterns in health and disease. Nutrients, 12: 334.
- Guine RP, Florença SG, Barroca MJ, Anjos O (2020) The link between the consumer and the innovations in food product development. Foods, 9: 1317.
- Ebert AW (2022) Sprouts and microgreens—novel food sources for healthy diets. Plants, 11: 571.
- Di Gioia F, Renna M, Santamaria P (2017) Sprouts, microgreens and “baby leaf” vegetables. Minimally processed refrigerated fruits and vegetables, 403-32.
- Aires A, Chapter 3-Brassica Composition and Food Processing. Processing and Impact on Active Components in Food; Preedy, V., Ed, 17-25.
- Maggioni L, von Bothmer R, Poulsen G, Branca F, Bagger Jørgensen R, (2014) Genetic diversity and population structure of leafy kale and Brassica rupestris Raf. in south Italy. Hereditas. 151: 145-58.
- Branca F, Di Bella MC, Arena D, Tribulato A, Kusznierewicz B, et al. (2022) Chemical characterization of wild populations of brassica oleracea complex Species (N= 9) for the content of their bioactive compounds. Acta Hortic.
- Treccarichi S, Ben Ammar H, Amari M, Cali R, Tribulato A, Branca F, (2023) Molecular markers for detecting inflorescence size of Brassica oleracea L. crops and B. oleracea complex species (n= 9) useful for breeding of broccoli (B. oleracea var. italica) and cauliflower (B. oleracea var. botrytis). Plants. 12: 407.
- Ben Ammar H, Sdouga D, Di Bella MC, Treccarichi S, Cali R, Rosa E, et al. (2021) Detection of glucosinolates metabolite pathway using SSR markers of Brassica oleracea complex species (n= 9) core collection. Acta Hortic. 1354: 105-10.
- Yan L, Zhou G, Shahzad K, Zhang H, Yu X, et al. (2023) Research progress on the utilization technology of broccoli stalk, leaf resources, and the mechanism of action of its bioactive substances. Front. Plant Sc. 14: 1138700.
- Picchi V, Lo Scalzo R, Tava A, Doria F, Argento S, et al. (2020) Phytochemical characterization and in vitro antioxidant properties of four Brassica wild species from Italy. Molecules, 25: 3495.
- Ben Ammar H, Picchi V, Arena D, Treccarichi S, Bianchi G, et al. (2022) Variation of Bio-Morphometric Traits and Antioxidant Compounds of Brassica oleracea L. Accessions in Relation to Drought Stress. Agronomy. 12: 2016.
- Arena D, Ben Ammar H, Rodriguez VM, Velasco P, Garcia G, et al. (2024) Exogenous Melatonin Affects the Morphometric Characteristics and Glucosinolates during the Initial Growth Stages of Broccoli. Agronomy, 14: 286.
- Barillari J, Canistro D, Paolini M, Ferroni F, Pedulli GF, et al. (2005) Direct antioxidant activity of purified glucoerucin, the dietary secondary metabolite contained in rocket (Eruca sativa Mill.) seeds and sprouts. J. Agric. Food Chem. 53: 2475-82.
- Sun-Ju Kim K, Gensho I, (2006) Glucosinolate profiles in the seeds, leaves and roots of rocket salad (Eruca sativa Mill.) and anti-oxidative activities of intact plant powder and purified 4-methoxyglucobrassicin, J. Soil Sci. Plant Nutr. 52: 394-400.
- Di Bella MC, Melilli MG, Treccarichi S, Tribulato A, Arena D, et al. (2022) Influence of irrigation regime on productive and qualitative traits of kale (Brassica oleracea var. acephala DC) under organic farming system. Acta Hortic. 1354: 301-8.
- Soengas P, Rodríguez VM, Velasco P, Cartea ME, (2018) Effect of temperature stress on antioxidant defenses in Brassica oleracea. Acs Omega, 3: 5237-43.
- Reshi ZA, Ahmad W, Lukatkin AS, Javed SB (2023) From nature to Lab: a review of secondary metabolite Biosynthetic pathways, Environmental influences, and in vitro approaches. Metabolites, 13: 895.
- He M, He CQ, Ding NZ, (2018) Abiotic stresses: general defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 9: 1771.
- Ben Ammar H, Arena D, Treccarichi S, Di Bella MC, Marghali S, et al. (2023) The Effect of Water Stress on the Glucosinolate Content and Profile: A Comparative Study on Roots and Leaves of Brassica oleracea L. Crops. Agronomy. 13: 579.
- Abbas K, Li J, Gong B, Lu Y, Wu X, Lü G, Gao H (2023) Drought Stress Tolerance in Vegetables: The Functional Role of Structural Features, Key Gene Pathways, and Exogenous Hormones. International Journal of Molecular Sciences, 24: 13876.
- Topcu Y, Dogan A, Sahin-Nadeem H, Polat E, Kasimoglu Z, Erkan M (2018) Morphological and biochemical responses of broccoli florets to supplemental ultraviolet-B illumination. Agriculture, Ecosystems & Environment, 259: 1-10.
- Liu Y, Singh SK, Pattanaik S, Wang H, Yuan L (2023) Light regulation of the biosynthesis of phenolics, terpenoids, and alkaloids in plants. Communications Biology, 6.
- Demmig‐Adams B, ADAMS III, WW (1992) Carotenoid composition in sun and shade leaves of plants with different life forms. Plant, Cell & Environment, 15: 411-9.
- Benzie IF, Strain JJ (1996) The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Analytical biochemistry, 239: 70-6.
- Brand-Williams W, Cuvelier ME, Berset C (1995) Use of a free radical method to evaluate antioxidant activity. LWT—Food Science and Technology, 28: 25-30.
- Sembiring EN, Elya B, Sauriasari R (2018) Phytochemical screening, total flavonoid and total phenolic content and antioxidant activity of different parts of Caesalpinia bonduc (L.) Roxb. Pharmacognosy journal, 10.
- De Pascale S, Arena C, Aronne G, De Micco V, Pannico A, Paradiso R, Rouphael Y (2021) Biology and crop production in Space environments: Challenges and opportunities. Life Sciences in Space Research, 29: 30-37.
- Benitez-Alfonso Y, Soanes BK, Zimba S, Sinanaj B, German L, et al. (2023) Enhancing climate change resilience in agricultural crops. Current biology, 33: R1246-61.
- Fritz MA, Rosa S, Sicard A (2018) Mechanisms underlying the environmentally induced plasticity of leaf morphology. Frontiers in genetics, 9: 478.
- Duan Y, Santiago FEM, Dos Reis AR, de Figueiredo MA, Zhou S et al. (2021) Genotypic variation of flavonols and antioxidant capacity in broccoli. Food Chemistry, 338: 127997.
- Amitrano C, Paglialunga G, Battistelli A, De Micco V, Del Bianco M et al. (2023) Defining growth requirements of microgreens in space cultivation via biomass production, morpho-anatomical and nutritional traits analysis. Frontiers in Plant Science, 14.
- Pattison PM, Tsao JY, Brainard GC, Bugbee B (2018) LEDs for photons, physiology and food. Nature, 563: 493-500.
- Huseby S, Koprivova A, Lee BR, Saha S, Mithen R et al. (2013) Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. Journal of Experimental Botany, 64.
- Pérez‐Balibrea S, Moreno DA, García‐Viguera C (2008) Influence of light on health‐promoting phytochemicals of broccoli sprouts. Journal of the Science of Food and Agriculture, 88: 904-10.
- Kuai J, Li X, Ji J, Li Z, Xie Yet al. (2022) Response of leaf carbon metabolism and dry matter accumulation to density and row spacing in two rapeseed (Brassica napus L.) genotypes with differing plant architectures. The Crop Journal, 10: 680-91.
- Roychowdhury R, Das SP, Gupta A, Parihar P, Chandrasekhar K et al. (2023) Multi-Omics Pipeline and Omics-Integration Approach to Decipher Plant’s Abiotic Stress Tolerance Responses. Genes, 14: 1281.
- Major N, Prekal jB, Perković J, Ban D, Užila Z et al. (2020) The Effect of Different Extraction Protocols on Brassica olerace a var. acephala Antioxidant Activity, Bioactive Compounds, and Sugar Profile. Plants, 9: 1792.
FIGURE 1
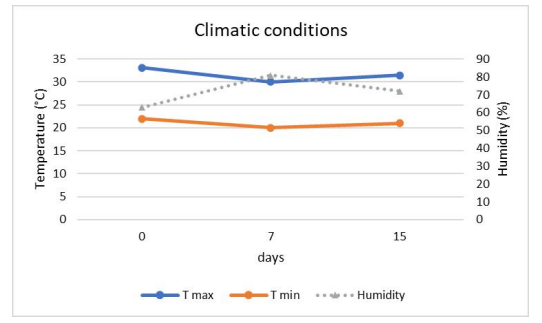
Figure 1: Greenhouse conditions during the growth cycle, maximum temperature (T max), minimum temperature (T min) and average relative humidity (%)
FIGURE 2
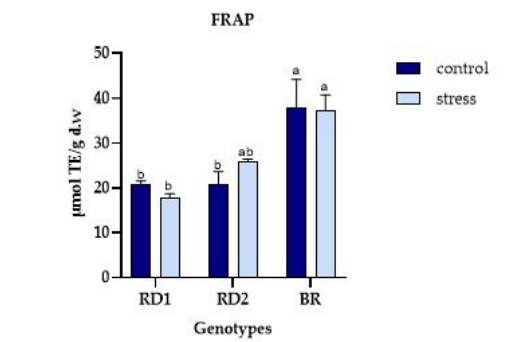
Figure 2: Variation of FRAP ( μmol TE/g d.w.) in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 3
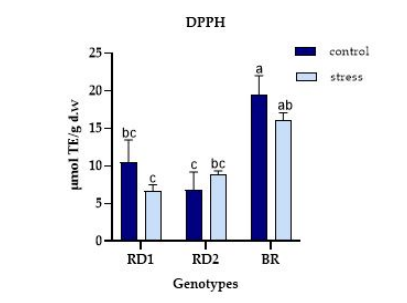
Figure 3: Variation of DPPH (μmol TE/g d.w.) in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 4
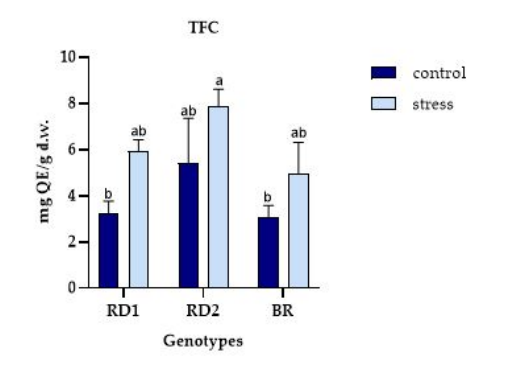
Figure 4: Variation of TFC (mg Quercetin Equivalents/g d.w.) in sprouts, microgreens, and baby leaves of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 5
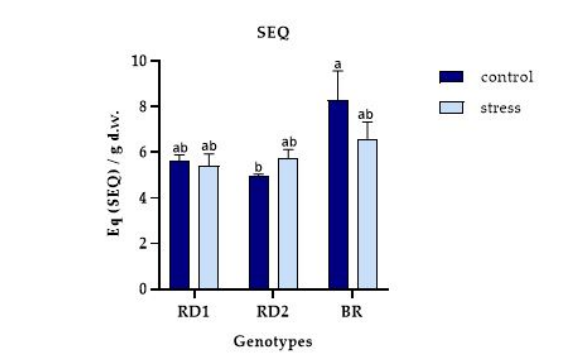
Figure 5: Total Glucosinolates (g SEQ/g d.w.) variation in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 6

Figure 6: Variation of the Total sugars (g/1000 g d.w.) in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 7
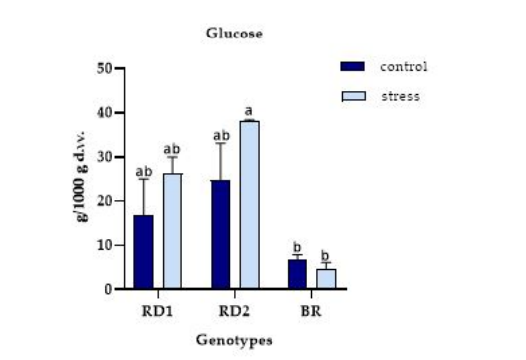
Figure 7: The variation of glucose (g/1000 g d.w.) in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 8
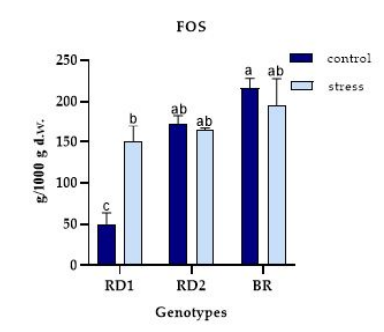
Figure 8: Variation of the FOS (g/1000 g d.w.) in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 9
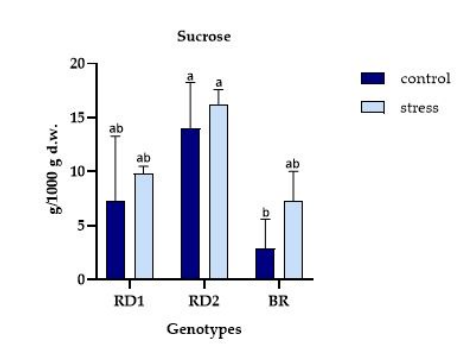
Figure 9: Variation of the sucrose (g/1000 g d.w.) in microgreens of Broccolo nero (BR), Eruca sativa var. darkita (RD1) and Eruca sativa var. lobata (RD2). Each value represents the mean ± S.E. of three replicates. The letters above bars indicate significant differences according to Tukey's test (p < 0.05)
FIGURE 10
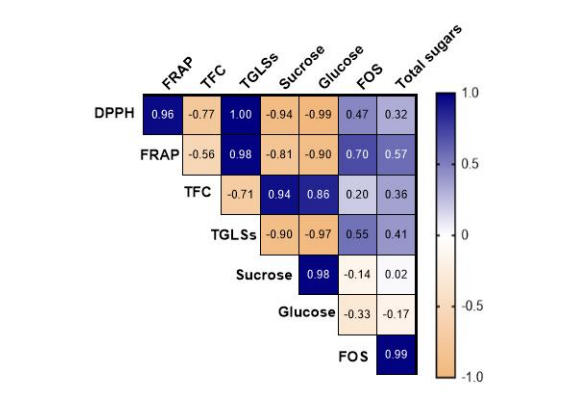
Figure 10: Pearson's correlation coefficients among the different variables studied for control condition
FIGURE 11
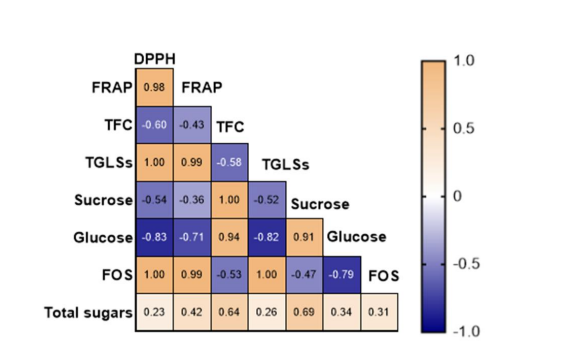
Figure 11: Pearson's correlation coefficients among the different variables studied for stress condition
FIGURE 12
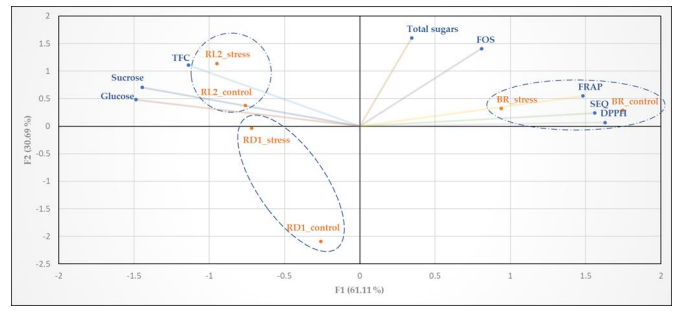
Figure 12: Principal component analysis biplot with control=SR100, stress= SR40, BR= Broccolo nero, RD1= Eruca sativa var. darkita and RD2= Eruca sativa var. lobata
Tables at a glance
Figures at a glance