Responses of Seedling Growth and Physiological Characteristics of Perennial Species Sophora alopecuroides L. to Warming and Nitrogen Deposition in Temperate Arid Habitats, China
Received Date: March 08, 2025 Accepted Date: April 08, 2025 Published Date: April 11, 2025
doi:10.17303/jacs.2025.4.103
Citation: Hong-Mei Zhao, Yu Zhao, Yi-Jun Bai, Xiao-Lin Zhang, Zhi-Yi Zeng, et al. (2025) Responses of Seedling Growth and Physiological Characteristics of Perennial Species Sophora alopecuroides L. to Warming and Nitrogen Deposition in Temperate Arid Habitats, China. J Adv Agron Crop Sci 4: 1-13
Abstract
The impacts of global climate change, particularly warming and nitrogen deposition, on plant growth and physiological traits have garnered significant attention in ecological research. This study investigates the responses of Sophora alopecuroides, a typical perennial plant in China's temperate arid region, to warming (W), nitrogen addition (N), and their combined effects (WN). The results reveal that W and WN treatments had no significant impact on seed germination rate, germination index, and germination potential, while N addition significantly inhibited seed germination rate. W and WN treatments significantly increased root - to - shoot ratio and plant height, whereas N addition significantly decreased these parameters. N addition significantly increased leaf biomass but reduced leaf number; W and WN treatments significantly promoted the accumulation of above - and below - ground biomass. Additionally, N addition increased leaf biomass ratio and root - to - shoot ratio, while WN treatment significantly increased stem and root biomass ratios. In terms of physiological metabolism, N addition significantly decreased chlorophyll, soluble sugar, and soluble protein content, while W treatment reduced malondialdehyde (MDA) content. WN treatment significantly increased nitrate reductase activity, and both W and N treatments strongly inhibited superoxide dismutase (SOD) activity. This study elucidates the adaptive mechanisms of S. alopecuroides to warming and nitrogen deposition, providing scientific evidence for understanding plant invasion processes under climate change and developing effective ecological management strategies.
Keywords: Biomass Accumulation; Climate Change; Nitrogen Deposition; Physicochemical Properties; Sophora Alopecuroides
Introduction
In recent years, global warming has become an irrefutable reality [30]. Moreover, as an important environmental factor, temperature exerts a profound influence on the growth and physiological characteristics of plants. Thus, global warming may have a significant impact on plant growth, development, reproduction, phenology, and migration and has become a hot topic in ecology and environmental science. Studies have shown that the effect of warming on plant growth may be beneficial [23], as well as inhibitory [28]. Warming can cause changes in plant traits, with a slower response in reproductivity [19], more pronounced and rapid response in plant growth [5]. However, the degree of its impact depends on the species and their environmental conditions. Studies have demonstrated that elevated temperatures enhance the germination rate, aboveground biomass, and reproductive capacity of Ipomoea cairica [26]. However, in some invasive plants such as Hypochaeris radicata L. and Leontodon taraxacoides L., warming has been shown to inhibit seed germination and seedling growth and even lead to vegetation degradation in northern Australia [28]. Climate warming may affect the competitive ability of alien invasive plants and enhance their physiological plasticity.
The Yili Grassland is an important part of the northern grasslands of China, and several important questions about the potential effects of warming need to be addressed: What are the effects of global warming on the composition and distribution of typical communities and the succession of toxic species communities in the grassland? How do the population life histories of these invasive toxic species respond to warming? What are the specific adaptive regulatory mechanisms in these processes? These are important theoretical issues of biology and ecology in a changing global environment and are also practical problems that need to be solved to control invasive plant species and ensure the security of grassland ecosystems and effective scientific management.
Global emissions of N have been increasing yearly since 2008, and as the demand for animal products increases, the emissions will gradually reach 135 Tg Nyr−1 by 2100 [9]. Studies have demonstrated that elevated nitrogen deposition can influence plant growth and allocation, alter community composition, and impact ecosystem functions [3]. Continuing nitrogen deposition will also alter nutrient cycling or interspecific competition among species [36] and may even promote the ecological invasion of certain species into new habitats [7]. Most studies have focused on forest and grassland ecosystems with high levels of nitrogen deposition, but relatively little attention has been given to arid and semiarid regions, especially in China [16, 33]. These areas are limited by water and nitrogen. Although water has a significant effect on the primary productivity of plants, nitrogen is the main factor when sufficient water is available [24]. Nitrogen deposition has been shown to have a significant effect on plant growth and community structure in the Chihuahuan Desert [2]. In the arid and semiarid regions of China, research on nitrogen addition has focused mainly on agro-ecosystems but significantly less attention has been given to the potential impacts of nitrogen deposition on natural ecosystems. Previous studies have indicated that nitrogen deposition might reduce biodiversity and alter the composition of herbaceous communities, thereby accelerating ecosystem degradation in desert grasslands [24]. Because of the extreme fragility and sensitivity of ecosystems in these areas, even small increases in nitrogen may have significant ecological effects, which would contribute a much higher proportion of production than in many high-productivity ecosystems [11]. Therefore, it is necessary to study the effects of increased nitrogen deposition on growth, production, and adaptation strategies in arid and semiarid regions under changing global environment. Such studies will contribute to a comprehensive understanding of the mechanisms by which nitrogen deposition affects invasive plants in arid and semiarid regions and also provide a scientific basis for grassland ecological conservation and management.
Sophora alopecuroides L. is a perennial herb or subshrub that grows mainly in western and central Asia. Because of its typical clonal characteristics, it propagates by both considerable amounts of seeds and underground rhizomes, which allows the species to invade and develop mono-dominant communities in farmland, temperate desert steppes, and temperate steppes. As a result, thousands of hectares of S. alopecuroides grassland have appeared in the Ili River Valley of China, which poses a serious threat to the local livestock husbandry, biodiversity, and ecosystem security. Therefore, the plant is recognized as a native invasive species in the arid areas of northwestern China [14, 20]. In recent years, how perennial species in temperate arid ecosystems respond to the synergistic effects of warming and nitrogen deposition have gradually attracted the attention of researchers globally, which is crucial to enhancing the understanding of the adaptation mechanisms of plants in changing global climates and environments. In previous studies, we found that S. alopecuroides has strong drought and stress tolerance [19], how it responds to new environmental stresses, such as warming and nitrogen deposition, remain unclear. Because it is the typical clonal and native invasive species of Leguminosae in the Yili grasslands and due to its toxicity, vicious invasion, and rapid expansion in arid and semiarid ecosystems, there is an urgent need to carry out studies on its life-history strategies at the population level in changing habitats, especially studies on how it responds to warming and nitrogen depletion. Therefore, in this study, which used S. alopecuroides as the research material, we attempted to answer the following two questions through simulated experiments of nitrogen deposition and warming: 1) How do clonal species S. alopecuroides cope with the synergistic effects of temperature increase and nitrogen deposition in temperate arid ecosystems? 2) What survival strategies and physiological regulatory mechanisms have been adopted by S. alopecuroides seedlings to adapt to elevated warming and nitrogen deposition? These findings will contribute to a deeper understanding of the ecological and physiological strategies used by invasive plants in the region to adapt to arid environments and will also provide some important insights that will enhance our understandings of the scientific prevention and control of various invasive species in these regions.
Materials and Methods
Materials
In mid-October 2022, mature pods of Sophora alopecuroides L. were collected from the suburbs of Yining City, Xinjiang, China, at the end of the growth period. The seeds were sorted, naturally air-dried, and stored in a 4 ℃ freezer. Before the experiment, uniformly sized and mature seeds were selected and sterilized using 0.01% HgCl2 and then soaked in distilled water for 48 h. The identification of S. alopecuroides L. was conducted by Prof. Yan Ping, and the herbarium specimen was deposited in the Institute of Botany, Chinese Academy of Sciences (specimen code: PE00214699). This species is widely distributed in northwest China, commonly found in mountainous grasslands and farmland, and is a local invasive species that can be obtained without the need for a permit at the time of sampling.
Methods
Potting treatment
In early October 2023, round and full S. alopecuroides seeds were selected, and the mechanical shell-breaking method was used to cut a small notch on the back of the seeds to avoid the inhibitory effect of the seed coat [18], which was completely immersed in a 10% sodium hypochlorite solution for 3 min to disinfect the seeds, and then taken out and rinsed repeatedly with distilled water for three to four times. The seeds were placed in distilled water for 24 h to fully absorb and set aside. The experiment was a factorial design with a total of four treatment groups, namely control (CK), warming (W), nitrogen application (N) and warming and nitrogen application interaction (WN). Each treatment group was planted in 10 pots, and 20 seeds of C. spinosa were uniformly sown in 21 cm × 15 cm × 18 cm pots with washed fine river sand, at a depth of 1 cm, with the same amount of sand in each pot, and a tray at the bottom of the pots.
Experimental design of treatment model
Based on the research method, the ambient temperature was set at 25°C (8:00-22:00) during the daytime and 18°C (22:00-8:00) at night; the temperature increase treatment was 28°C (8:00-22:00) during the daytime and 21°C (22:00-8:00) at night, with a light intensity of 6,000 LX. The control temperature was set as the temperature of Yining City for 30 years (1989 ~ 2019) monthly average temperature from March to June, 25°C during the day and 18°C at night. The temperature increase treatment was to increase the temperature by 3°C on the basis of the average temperature of the control group for the experimental design. The simulated nitrogen deposition was set at two levels, 0 and 12.5 mmol/L, and the total applied nitrogen of the nitrogen treatments was equivalent to the annual nitrogen deposition near the Gurbantunggut Desert (~1 g N·m-2·a-1) [35] and the maximum nitrogen deposition in the Mojave Desert of the United States (~4.03 g N·m-2·a-1) [4]. A 0.5-fold Ms nutrient solution (Macroelement, Microelement and Iron element) was prepared and watered at 7-day intervals, with approximately 200 mL of solution per potted plant per spray, each time at 10 am. Seedlings from each treatment group were harvested after waiting for the cotyledons to fall off naturally. The experimental design is shown in Figure 1.
Indicator measurement
Growth indicators
During the experiment, the seedlings were dug up with roots from 30 healthy Sophora alopecuroides plants and the sand was treated with a fine brush. Plant height and main root length of S. alopecuroides seedlings were measured using a meter scale with an accuracy of 1 mm. The basal and root diameters of the seedlings were measured using vernier calipers with an accuracy of 0.01 mm. Roots, stems and leaves were separated and weighed using an electronic balance with an accuracy of 0.001 g. The fresh weights of roots, stems and leaves were used for calculating the growth traits such as the leaf biomass ratio, the root biomass ratio, the stem biomass ratio, and the root-crown ratio. The number of compound leaves per plant was counted and the leaves were scanned using EPSON leaf area scanner and morphologically characterized using Image J image analysis software to obtain leaf area data in 30 replicates.
Seed germination
The potted plants were incubated in an incubator and observed to record the daily germination. The radicle breaking through the seed coat was used as a sign of germination [22], the germination potential was recorded statistically from the 3rd day, the germination rate was recorded on the 7th day [13], and the seed germination experiment ended on the 10th day.
Gt denotes the total number of germinations on day t, while Dt represents the corresponding number of days of germination. G6 denotes the total number of germinations on day 6.
Physicochemical indicators
Four healthy S. alopecuroides seed plants were randomly selected for each treatment, and mature leaves were selected from each plant, which were quickly put into the ice box after picking, and 0.1g of fresh leaves were weighed for the determination of physiological and biochemical indexes. SOD activity was measured using SOD superoxide dismutase kit, NR activity was measured using NR nitrate reductase kit, malondialdehyde content was measured using the thiobarbituric acid reaction, and the protein content was measured using the coomassie brilliant blue staining G250 method [12]. thiobarbituric acid reaction colorimetric method [1], and soluble sugar content was determined by anthrone method [8], with 4 sets of replicates.
Statistical analysis
Excel 2010 software was used for preliminary data collation, and SPSS (version 26.0) was used for data analysis to compare the significance of differences between different treatments of the same indicator by two-way analysis of variance (ANOVA). Graphs were plotted using Origin 2021 software.
Results and Analysis
Effects of nitrogen application, warming, and their interaction on S. alopecuroides seed germination
As shown in Figure. 2, for S. alopecuroides seeds, in comparison to the control group, nitrogen application alone resulted in significant decreases in germination rate, germination force, and germination index of 13%, 16.5%, and 20.6%, respectively (Figure 2A, B and C). However, there were no significant differences in germination rate, germination force, and germination index compared to the control group under either the warming treatment alone or the combined treatment of warming and nitrogen application (Figure 2A, B and C). This indicates that nitrogen application negatively affects the germination performance of S. alopecuroides seeds, whereas warming treatment alone or in combination with nitrogen application has no significant effect on germination.
Effects of nitrogen application, warming, and their combination on the growth of S. alopecuroides seedlings
In this study, following the application of the different treatments to S. alopecuroides, several changes in root diameter, stem diameter, plant height, and primary root length were observed Table 1. Compared to the control group, nitrogen application resulted in significant reductions in stem diameter, plant height, and primary root length of 33.3%, 19.4%, and 24%, respectively. However, root diameter exhibited no significant change. In contrast, under the interaction of warming and nitrogen application, stem diameter and plant height increased significantly compared to the control group (Table 1). Under the combined treatment of warming and nitrogen application, root diameter exhibited a 7.2% and 28.5% increase, stem diameter exhibited a 23.6% and 19% increase, plant height exhibited a 20.3% and 13.2% increase, and primary root length exhibited a 19.5% and 13.8% increase, respectively. A comparison of the different treatment groups revealed that warming could promote to varying degrees increases in stem diameter, plant height, and primary root length of S. alopecuroides seedlings, whereas nitrogen application could inhibit these effects to varying degrees.
Effects of nitrogen application, warming, and their combination on biomass accumulation and allocation in S. alopecuroides
Table 2 indicates that the combined treatment of warming and nitrogen application significantly increased the root biomass of seedlings by 257.7% and 351.3%, respectively. Furthermore, the leaf area increased by 17.9% and 11.4%, leaf biomass increased by 70.2% and 56.2 %, and stem biomass increased by 114.9% and 92 %, respectively (Table 2). However, in the independent nitrogen application treatment, there were no significant changes in root biomass and stem biomass compared to the control group. in contrast, leaves exhibited 87.0% increase in biomass of, but leaf area and leaf number significantly decreased (Table 2). These findings indicated that S. alopecuroides seedlings exhibited a positive response in root growth and leaf development under warming and the combined treatment of warming and nitrogen application, indicating both promoting and interaction effects.
Figure 3 illustrates that sole nitrogen application treatment showed no significant changes in root biomass ratio and stem biomass ratio of seedlings in compared to the control group (Figure 3A, and B). However, the leaf biomass ratio and root-to-shoot ratio increased significantly (Figure 3C, and D). This indicates that nitrogen application can promote photosynthesis and nutrient absorption in S. alopecuroides seedlings, resulting in improved efficiency of leaf tissue and spatial utilization. Under the warming treatment alone, the root biomass ratio and stem biomass ratio increased significantly (Figure 3A, and B) compared to the control group, whereas, the leaf biomass ratio and root-to-shoot ratio did not show significant changes (Figure 3C, and D). This indicated that the warming treatment could enhance plant root growth and increase plant biomass. Under warming treatment alone, root biomass ratio and stem biomass ratio of S. alopecuroides seedlings significantly increased by 143% and 46.8%, respectively (Figure 3A, and B), while leaf biomass ratio and root-to-shoot ratio did not change significantly (Figure 3C, and D) as compared to control. These findings indicate that the combined treatment of warming and nitrogen application promotes plant growth and significantly increases biomass accumulation in both roots and above ground stems.
Effects of nitrogen application, warming, and their combination on physicochemical indicators of S. alopecuroides
According to the results in Figure. 4, nitrogen application treatment significantly reduced the soluble sugar content of S. alopecuroides seedlings by 21.2% compared to the control (Figure 4B). However, no significant changes were observed in protein, chlorophyll, and MDA contents (Figure 4A, C, and D). However, the warming treatment resulted in a significant increase of 15.7% in soluble sugar content, while protein, chlorophyll, and MDA contents did not show significant changes. The combined treatment of warming and nitrogen application resulted in a significant decrease of 19% and 22.8% in the protein and chlorophyll contents of S. alopecuroides seedlings compared to the control group (Figure 4C, and D). However, no significant differences were observed in soluble sugar content or MDA content. These findings suggest that nitrogen application, warming, and their combined effects have a limited impact on the biochemical indicators of S. alopecuroides seedlings.
The enzyme activities of NR and SOD in S. alopecuroides seedlings exhibited significant differences among the different treatment groups (Figure 5A and B). Nitrate reductase activity was significantly higher than 56% of control, 37.4% of warming treatment and 20.4% of nitrogen application under combined treatment of warming and nitrogen application, respectively (Figure 5A). Compared to the control group, both the sole nitrogen application and sole warming treatments demonstrated a significant increase in NR activity (Figure 5A). The combined treatment of warming and nitrogen application exhibited the greatest increase in NR activity, followed by sole nitrogen application and sole warming treatments. Concerning SOD, all treatment groups exhibited a significant reduction in enzyme activity compared to the control group. The control group exhibited the highest SOD enzyme activity, followed by the sole nitrogen application treatment, and the combined treatments of warming and nitrogen application showed the lowest activity (Figure 5B). The combined treatments of warming and nitrogen application significantly increased NR activity in S. alopecuroides seedlings, while exerting a substantial inhibitory effect on SOD activity. The sole nitrogen application and sole warming application also significantly affected enzyme activity, although to a lesser extent than the combined application of warming and nitrogen.
Discussion
The main findings of the study showed that climate warming significantly boosted growth parameters of invasive plants, such as biomass, stem length and leaf area, thereby enhancing their competitive advantage. For example, S. alopecuroides seedlings showed a significant increase in biomass, height, leaf area, and number of true leaves under warm treatments, which is consistent with previous findings [23,26,27] In addition, antioxidant enzyme activities increased under warm conditions, suggesting that the antioxidant defense system of plants is effective in mitigating the damaging effects of heat-induced reactive oxygen species [21]. The effects of nitrogen addition on plant growth are complex and variable, with some plants exhibiting growth promotion under moderate nitrogen addition, while others may be inhibited. For example, N addition significantly increased biomass allocation and growth parameters of Solidago canadensis and coarse-fruited commiphora [25], but at the same time may also negatively affect the stability and positive succession of desert ecosystems [24]. In this study, N addition significantly reduced the germination rate of S. alopecuroides seeds as well as stem diameter, primary root length and plant height of seedlings, while leading to a decrease in chlorophyll and soluble sugar content, suggesting that nitrogen deposition has an inhibitory effect on plant growth and physiological status.
Combined with the existing literature, these results may be attributed to the fact that N deposition alters soil N availability, which in turn affects plant biomass distribution patterns and growth strategies [2,32]. Increased NR activity by N addition may have accelerated the nitrate reduction process and reduced nitrate accumulation, thus affecting plant growth [34].Decreased SOD activity, on the other hand, may indicate that plants are challenged with oxidative stress despite increased antioxidant enzyme activity, which may be a reason for the inhibition of plant growth under N addition. The main findings of this study suggest that moderate nitrogen deposition can promote above-ground plant growth within a certain range and change the distribution pattern of plant biomass, which in turn affects seedling growth [32]. However, there are differences in the response of different plants to nitrogen addition, with some plants showing significant biomass increases under high nitrogen conditions, such as Bouteloua eriopoda and Larrea tridentata [2], while S. alopecuroides showed a significant inhibitory effect. It was shown that nitrogen deposition reduced the germination rate of S. alopecuroides seeds as well as the stem diameter, primary root length and height of the plant, while significantly reducing chlorophyll and soluble sugar content, decreasing SOD activity while increasing NR activity, and significantly increasing leaf biomass ratio and root-crown ratio.
The existing literature suggests that the mechanism of nitrogen deposition on plant growth is complex and involves the regulation of nitrogen metabolism, photosynthesis efficiency and antioxidant system [6,15,29,31]. Nitrogen excess may lead to an imbalance of nitrogen metabolism in plants, affecting photosynthetic capacity and antioxidant defense system, which ultimately manifests as growth inhibition [10]. In addition, nitrogen deposition may indirectly affect the nutrient uptake and utilization efficiency of plants by changing the structure of soil microbial communities [17]. However, there are some limitations in this study. First, the experiment focused only on a single invasive plant, S. alopecuroides, and failed to comprehensively reflect the integrated response of multiple plants to climate change and N addition. Second, the study did not explore differences in the effects of different temperature increases on plant growth, which may affect the generalizability of the results. Future studies should consider expanding the range of plant species and delving into differences in the effects of different diurnal warming on plant physiology and growth, and how these effects vary over time. In addition, research needs to further explore the long-term effects of climate change and nitrogen deposition interactions on plant populations and ecosystem structure and function.
Conclusions
Based on the study, S. alopecuroides seedlings have developed several survival strategies and physiological mechanisms to adapt to warming and nitrogen deposition. Warming significantly enhances growth and biomass accumulation, especially in aboveground biomass. Although nitrogen deposition negatively impacts seed germination, plant height, stem diameter, and physiological features like soluble sugars and SOD, these seedlings still show increased aboveground biomass in elevated temperatures. The combined effects of warming and nitrogen result in continued growth but also inhibit certain physiological processes, such as soluble proteins and chlorophyll. Overall, temperature is a crucial factor for promoting growth, and the interplay between temperature and nitrogen plays a significant role in the ecological and physiological responses of these perennial plants in temperate arid ecosystems. This research offers valuable insights into plant survival strategies under global climate change and informs future conservation efforts in desert ecosystems.
Acknowledgments
We acknowledge the Open Innovation Lab of History, Geography and Tourism School, Shangrao Normal University for providing the experimental conditions and lab analysis.
Author Contributions
Hongmei Zhao, Xiaolin Zhang, Zhiyi Zeng, and Zhijie Wangyu contributed to the components of the experiments and data collection. Data analysis was performed by Yu Zhao and Yijun Bai. The experimental design was carried out by Tiancui Shang and Ying Liu. The first draft of the manuscript was written by Hongmei Zhao, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31360101;32360265), the Science and Technology Foundation of Jiangxi Educational Committee (201701,GJJ211718), and the Open Project of Key Laboratory of Microbial Resource Protection, Development and Utilization (YLUKLM202003).
Data Availability
Data will be made available on request.
Declarations
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
- Andre DdAN, Jose TP, Joaquim Ea-F, Carlos EBdA, Ene aG-F (2006) Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environmental and Experimental Botany, 56: 87-94.
- Báez S, Fargione J, Moore DI, Collins SL, Gosz JR (2007) Atmospheric nitrogen deposition in the northern Chihuahuan desert: Temporal trends and potential consequences. Journal of Arid Environments, 68: 640-51.
- Broadbent A, Stevens CJ, Peltzer DA, Ostle NJ, Orwin KH (2017) Belowground competition drives invasive plant impact on native species regardless of nitrogen availability. Oecologia, 186: 577-87.
- Brooks ML (2003) Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. Journal of Applied Ecology, 40: 344-53.
- Cavieres LA, Sanhueza AK, Torres-Mellado G, Casanova-Katny A (2017) Competition between native Antarctic vascular plants and invasive Poa annua changes with temperature and soil nitrogen availability. Biological Invasions, 20: 1597-610.
- Chen XT, Kameshwar AKS, Chio C, Lu F, Qin WS (2019) Effect of KNO3 on Lipid Synthesis and CaCO3 Accumulation in Pleurochrysis dentata Coccoliths with a Special Focus on Morphological Characters of Coccolithophores. International Journal of Biological Sciences, 15: 2844-58.
- Dawson W, Schrama M, Austin A (2016) Identifying the role of soil microbes in plant invasions. Journal of Ecology, 104: 1211-8.
- Dong CN, Peng XX, Yang XN, Wang CG, Yuan LY, et al (2024) Physiological and Transcriptomic Responses of Bok Choy to Heat Stress. Plants, 13: 1093-110.
- Fowler D, Steadman CE, Stevenson D, Coyle M, Rees RM, et al. (2015) Effects of global change during the 21st century on the nitrogen cycle. Atmospheric Chemistry and Physics, 15: 13849-93.
- Genchi G, Sinicropi MS, Lauria G, Carocci A, Catalano A (2020) The Effects of Cadmium Toxicity. International Journal of Environmental Research and Public Health, 17.
- Harpole, Stanley W, Tilman, David (2007) Grassland species loss resulting from reduced niche dimension. Nature, 446: 791-3.
- Hu Y, Zuo XA, Yue P, Zhao SL, Guo XX, et al. (2020) Increased Precipitation Shapes Relationship between Biochemical and Functional Traits of Stipa glareosa in Grass-Dominated Rather than Shrub-Dominated Community in a Desert Steppe. Plants, 9.
- Lai LM, Chen LJ, Zheng MQ, Jiang L, Zhou JH, et al (2019) Seed germination and seedling growth of five desert plants and their relevance to vegetation restoration. Ecology and Evolution, 9: 2160-70.
- Lei LJ, Zhao Y, Shi K, Liu Y, Hu YX, Shao H (2021) Phytotoxic Activity of Alkaloids in the Desert Plant Sophora alopecuroides. Toxins, 13.
- Li H, Zhang Y, Li H, V. P. Reddy G, Li Z, et al (2023) The nitrogen-dependent GABA pathway of tomato provides resistance to a globally invasive fruit fly. Frontiers in Plant Science, 14.
- Li T, Cui LZ, Liu LL, Wang H, Dong JF, et al (2022) Characteristics of nitrogen deposition research within grassland ecosystems globally and its insight from grassland microbial community changes in China. Frontiers in Plant Science, 13.
- Liu Y, Yang HC, Liu Q, Zhao XW, Xie S, et al (2021) Effect of Two Different Sugarcane Cultivars on Rhizosphere Bacterial Communities of Sugarcane and Soybean Upon Intercropping. Frontiers in Microbiology, 11.
- Liu Y, Zhao Y, Cui D, Leng JM, Dong FH (2017) Effect of cotyledon damage on the early growth of Sophora alopecuroides seedling. Acta Pratacul Turae Sinica. 26: 139-45.
- Liu YZ, Reich PB, Li GY, Sun SC (2011) Shifting phenology and abundance under experimental warming alters trophic relationships and plant reproductive capacity. Ecology, 92: 1201-7.
- Lv DK, Ba Ys, Zhao Y, Ou YY (2011) Seed Germination Characteristics of Sophora alopecuroides L. Journal of Grassland Science, 19: 889-92.
- MÁJEKOVÁ M, FRANCESCO DB, JIŘÍ D, JAN L (2014) Plant functional traits as determinants of population stability. Ecology, 95: 2369-74.
- Miransari M, Smith DL (2014) Plant hormones and seed germination. Environmental and Experimental Botany, 99: 110-21.
- Piper, Frida I, Fajardo, Alex, Cavieres, Lohengrin A (2013) Simulated warming does not impair seedling survival and growth of Nothofagus pumilio in the southern Andes. Perspectives in Plant Ecology, Evolution and Systematics, 15: 97-105.
- Su JQ, Liu XR, Hui R, Zhao Y, Liu YM (2012) The Effect of Nitrogen Deposition on Species Diversity and Community Composition of Herbaceous Vegetation in Desert Steppe. The Northwestern Botanical Journal, 32: 97-105.
- Wan LY, Qi SS, Zou CB, Dai ZC, Ren GQ, et al. (2019) Elevated nitrogen deposition may advance invasive weed, Solidago canadensis, in calcareous soils. Journal of Plant Ecology, 12: 846-56.
- Wang RL, Zeng RS, Peng SL, Chen BM, Liang XT, Xin XW (2011) Elevated temperature may accelerate invasive expansion of the liana plant (Ipomoea cairica). Weed Research, 51: 574-80.
- Wang ZX, He ZS, He WM (2021) Nighttime climate warming enhances inhibitory effects of atmospheric nitrogen deposition on the success of invasive Solidago canadensis. Climatic Change, 167.
- Williams AL, Wills KE, Janes JK, Jacqueline K, Vander S, et al. (2007) Warming and free‐air CO2 enrichment alter demographics in four co-occurring grassland species. New Phytologist, 176: 365-74.
- Xia AT, Wu YY, Xiang JQ, Yin HQ, Ming JJ, Qin ZH (2023) Quantification of Glucose Metabolism and Nitrogen Utilization in Two Brassicaceae Species under Bicarbonate and Variable Ammonium Soil Conditions. Plants, 12: 3095-118.
- Xue HH, Shi F, Gennaretti F, Fu YS, He B, et al (2023) Evidence of advancing spring xylem phenology in Chinese forests under global warming. Science China Earth Sciences, 66: 2187-99.
- Yang W, Zhou Z, Chu ZH (2023) Emerging Roles of Salicylic Acid in Plant Saline Stress Tolerance. International Journal of Molecular Sciences, 24: 3388-408.
- Zhang J, Zuo X, Zhao X, Ma J, Medina-Roldan E (2020) Effects of rainfall manipulation and nitrogen addition on plant biomass allocation in a semiarid sandy grassland. Scientific Reports, 10.
- Zhang T, Liu MZ, Huang XD, Hu W, Qiao N, et al. (2020) Direct effects of nitrogen addition on seed germination of eight semi‐arid grassland species. Ecology and Evolution, 10: 8793-800.
- Zhang T, Yang SB, Guo R, Guo JX (2016) Warming and Nitrogen Addition Alter Photosynthetic Pigments, Sugars and Nutrients in a Temperate Meadow Ecosystem Plos One, 11: 1932-6203.
- Zhang Y, Zheng LX, Liu XJ, Jickells T, Neil Cape J, et al (2008) Evidence for organic N deposition and its anthropogenic sources in China. Atmospheric Environment, 42: 1035-41.
- Zhu JX, Chen Z, Wang QF, Xu L, He NP, et al. (2020) Potential transition in the effects of atmospheric nitrogen deposition in China. Environmental Pollution, 258: 113739.
FIGURE 1
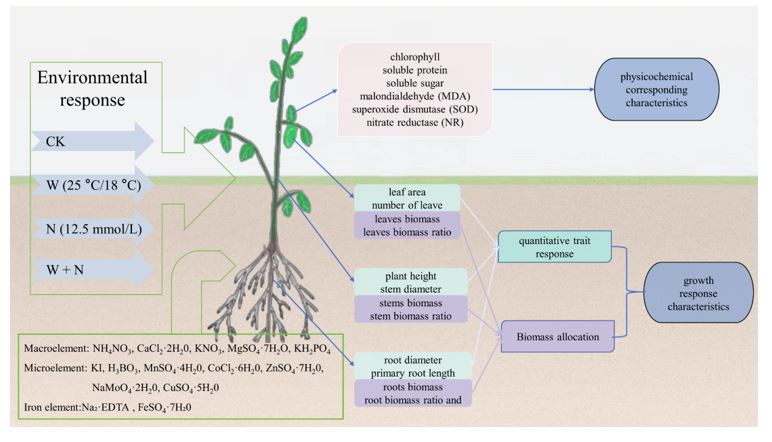
Figure 1: Depicts the experimental layout. S. alopecuroides were cultivated in MS nutrient solution for cotyledon shedding. Throughout the growth period, various treatments were applied, including a control, warming, nitrogen addition, and the interaction of warming and nitrogen addition.
FIGURE 2
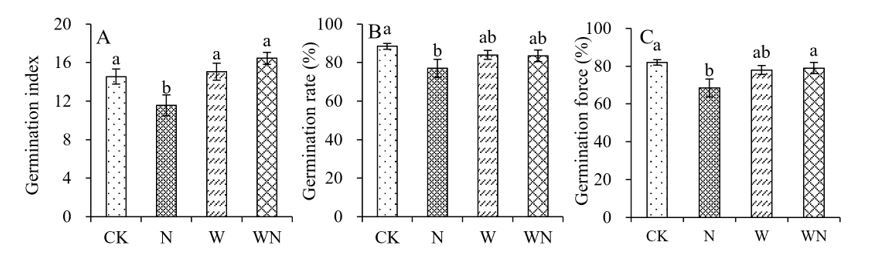
Figure 2: Impacts of nitrogen application, warming, and their interaction on the germination of S. alopecuroides. A: Germination index, B: Germination rate, C: Germination force
FIGURE 3
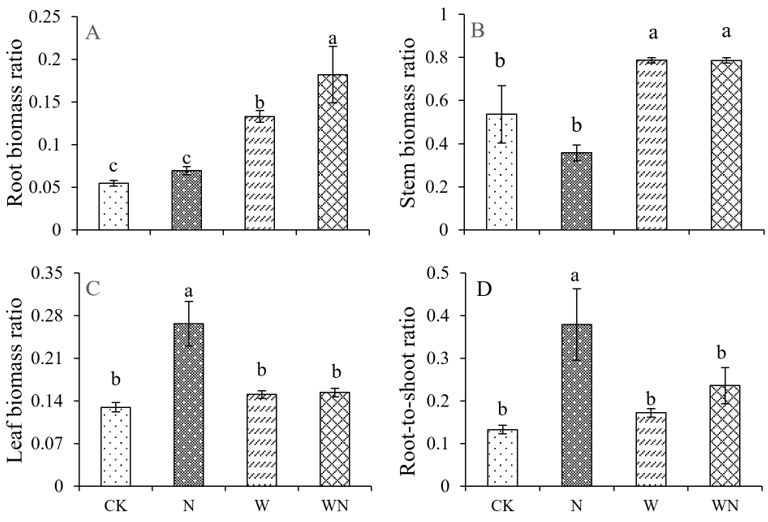
Figure 3: Effects of nitrogen and warming treatments on biomass allocation in S. alopecuroides seedlings. A: Root biomass ratio, B: Stem biomass ratio, C: Leaf biomass ratio, D: Root-to-shoot ratio
FIGURE 4
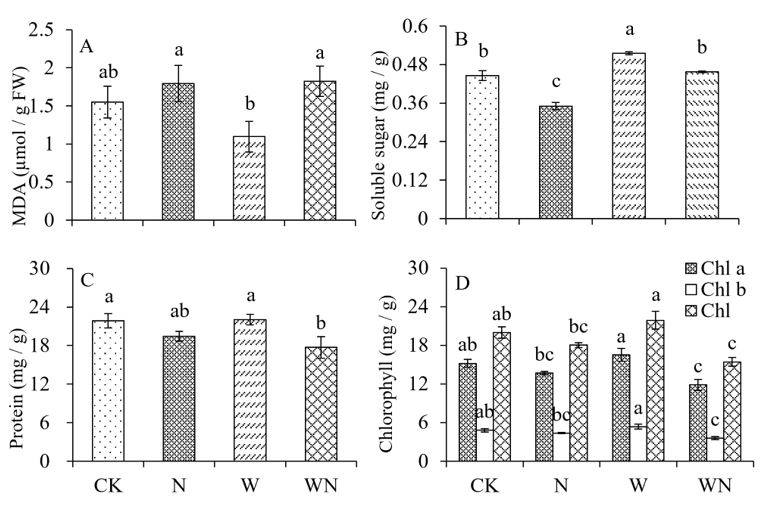
Figure 4: Effects of nitrogen and warming treatment on physicochemical indices of S. alopecuroides seedlings. A: MDA, B: Soluble sugar content, C: Protein content, D: chlorophyll content
FIGURE 5
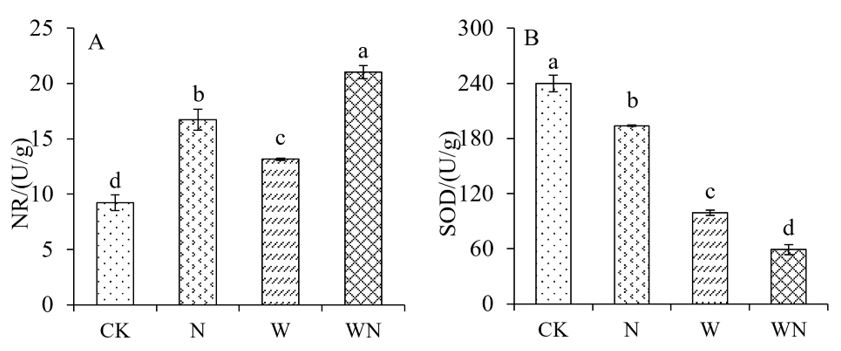
Figure 5: Effects of nitrogen and warming treatments on enzyme activities of S. alopecuroides seedlings. NR: Nitrate Reductase, SOD: Superoxide Dismutase, A: NR, B: SOD
Tables at a glance
Figures at a glance