“In-Vitro” Expression Profile of HSP 70.1 And HSP 90.1 Genes in Peripheral Blood Mononuclear Cells of Indian Native Tharparkar and Crossbred Vrindavani Cattle
Received Date: February 05, 2022 Accepted Date: March 05, 2022 Published Date: March 08, 2022
doi: 10.17303/javm.2022.1.102
Citation: Lakshya Veer Singh, G Ravi Kumar, Shishir Kumar Gupta, Smita Gupta, Pragya Singh et al. (2022) “In-Vitro” Expression Profile of HSP 70.1 And HSP 90.1 Genes in Peripheral Blood Mononuclear Cells of Indian Native Tharparkar and Crossbred Vrindavani Cattle. J Anim Biol Vet Sci 1: 1-10.
Abstract
Circulating peripheral blood mononuclear cells (PBMCs) can be used as an effective model to understand the heat stress response of different cattle types and buffaloes. Environmental temperature is one of the significant abiotic influences that impact the normal biological function and productive performance of dairy cattle. This investigation was undertaken to determine the temporal profile of HSPs (HSP70.1 and HSP90.1) expression in circulating PBMCs isolated from Tharparkar and crossbred (Vrindavani) cattle in response to lethal heat shocks at T40 °C, T41 °C and T42 °C for 13 hours. The quantitative real-time reverse transcription (qRT-PCR) expression data showed significant increase in mRNA levels of the both HSPs genes in heat stressed PBMCs. Out of two HSPs, HSP 90.1 was relatively more expressed followed by HSP70.1 in these two cattle types, indicating its prominent role as molecular chaperone to stabilize the native conformation of proteins. Also, the response elicited was different for both the cattle as the level of expression of HSPs throughout the time period of heat stress was higher in crossbred cattle than in Tharparkar. Results indicated that PBMCs from crossbred Vrindavani cattle are less tolerant to heat exposure than those from Indian native Tharparkar cattle, which is associated with higher expression levels of HSP70.1 and HSP90.1 genes.
Keywords: PBMCs; Heat stress; HSP; Tharparkar; Crossbred; Vrindavani
Introduction
India has the largest livestock population in the world and ranks first in respect of cattle and buffalo population. Cattle constitute 37.5% of its total livestock population (BAHS Basic Animal Husbandry Statistics 2012) [1] and contribute around 50% of milk production in India (Rajoriya et al. 2014 and Sharma et al. 2013; Sengar et al. 2018) [2-4]. Indigenous breeds of cattle are well known for their thermotolerant capability and high disease resistance as well as major milk producing species in the Indian subcontinent. Indian zebu cattle (Bos indicus) breeds are well known for their adaptation to different agro-climatic conditions, their hardiness and survival under stressful conditions. As these cattle are thermotolerant, therefore, the these cattle have significantly less impact of heat stress on growth rate, milk yield as well as on reproductive functions (Hansen 2004; O’Bannon et al. 1955; Johnson 1965; Johnston et al. 1963; Chen and Qiu 2012) [5-9]. Among these cattle breeds, Indian native cattle breeds viz. Ongole, Sahiwal, Tharparkar etc. show much better heat tolerance than Bos Taurus and crossbred cattle (Rajoriya et al. 2014; Singh et al. 2014) [2,10].
Tharparkar is one of the most important dual-purpose breeds and are very resistant to many tropical diseases and have good heat tolerance ability (Rajoriya et al. 2014; Deb et al. 2013) [2,11]. During past few decades, due to crossbreeding program and mechanization of agriculture in the country, various indigenous cattle breeds which form the core of traditional agriculture are being diluted. Currently, there is around 22.1 million crossbred cattle population including Holstein Friesian (HF), Jersey crosses and Vrindavani in India (Narayan et al. 2007) [12]. Though the crossbred animals are known for high production, they are also found to be more susceptible to climatic stress conditions (Narayan et al. 2007; Deb et al. 2014; Driver and Khatib 2013) [12-14]
Heat shock proteins (HSPs) are the conserved protein and play an important role in protein folding and unfolding as molecular chaperones and in protection of cells from various stresses including heat stress (Mathew and Morimoto 1998; Li and Shrivastava 2004; Sodhi et al. 2013) [15-17]. HSPs are present in cells under normal conditions, but when cells are exposed to a higher temperature, the expressions of these proteins is up-regulated and play an important role in protein stabilisation and folding of denatured proteins in response to heat shock stress. Therefore, differential expression of HSPs under thermal stress may partially explain the relative stress tolerance of native breeds of cattle compared to exotic breeds (West 2003; Wheelock et al. 2010; Hu et al. 2016) [18-20]. In tropical, subtropical, and arid regions, high ambient temperature is one of the major factors affecting high producing animal production (Gill et al. 2017) [21]. Heat tolerance can be defined as “ability of the body to endure the impact of a hot environment without suffering ill-effects” (Bianca 1961) [22].
Heat shock proteins (HSPs) have been classified into five families (100, 90, 70, 60 and small HSPs) according to their molecular weight (Kristensen et al. 2004; Kumar et al. 2015; Verma et al. 2016) [23-25]. These are a group of highly conserved proteins which are expressed by all living organisms. HSPs are components of anti-stress mechanisms and allow cells to adapt to gradual environment changes. These molecular chaperones encompass several families in which HSP70 and 90 are the most temperature sensitive and highly conserved of the HSPs. HSP70 and HSP90 proteins protect cells from thermal or oxidative stress by temporarily binding to hydrophobic residues exposed by stress, these HSPs prevents these partially denatured proteins from aggregating, and allows them to refold (Kregel 2002; Picard 2002; Deb et al. 2014; Kumar et al. 2015) [26,27,13,24].
The objective of the present study was to characterise the stress response of PBMCs from Indigenous (Tharparkar) and crossbred cattle (Vrindavani) to heat challenges of varying temperatures and exposure times based on the expression profile of HSP70.1 and HSP90.1 genes.
Materials and Methods
Experimental animals
The present study was conducted on Tharparkar (Bos indicus) and Vrindavani crossbred cow, maintained at cattle and buffalo farm, Indian Veterinary Research Institute, Izatnagar. Vrindavani are crossbred cattle, which have recently been developed, and have the exotic inheritance of Holstein-Friesian, Brown Swiss, Jersey and indigenous inheritance of Hariana cattle (Singh et al. 2011) [28]. About 8-10 ml of venous blood was collected from each animal (15 animals of each breed) under sterile conditions from jugular vein in vacutainer containing EDTA.
Isolation of PBMCs
The PBMC were isolated by density gradient centrifugation (Singh et al. 2014). Briefly, blood was diluted, layered over Ficoll-Histopaque-1077 (Sigma, MO, USA), and centrifuged. The mononuclear cell band was recovered and washed twice in phosphate buffer saline (PBS). Residual red blood cells were eliminated by hypotonic shock treatment using redistilled water. The PBMC recovery and viability were determined cell counter slide (Countess, Invitrogen, USA) and cell number was counted in a cell counter (Countess, Invitrogen, USA) count using the trypan blue exclusion method. Viability of PBMC typically exceeded 80% both in Tharparkar cattle and Vrindavani cattle. The PBMC were resuspended at 1×106 viable cells/mL in Roswell Park Memorial Institute 1640 medium (RPMI 1640 medium) (HiMedia, India) containing 25 mM HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) (HiMedia, India) supplemented with 10% heat-inactivated fetal bovine serum, 100 U of penicillin, 100 μg of streptomycin, and 0.25 μg of amphotericin B/mL (HiMedia, India). The time between blood collections and establishment of cultures was less than 6 h.
Heat treatment
The PBMC isolated from the 15 Tharparkar and Vrindavani cattle were subjected to heat shock treatment. Cells were divided into 4 samples viz. control (T1), T2, T3, T4 each receiving different heat treatments. The 65-h incubation represents an optimal time to guarantee cell viability and elicit maximum proliferative response in bovine PBMC (Lacetera et al. 2006) [29]. Control cells were exposed to 37°C continuously for 65 h or while treatments cells were kept at 40, 41, or 42 °C, respectively, for 65 h. Treatment at 37 °C was adopted to mimic normothermia; 40, 41 and 42 °C groups mimicked conditions of severe hyperthermia.
PBMC cultures for Proliferation assay
The PBMC (100 μL) were added into quadruplicate wells of 24- well, flat-bottomed tissue culture plates and each samples put in four times. Tissue culture plates were subjected to the treatment protocol in an atmosphere of 95% air and 5% CO2. After the first 24 h of incubation, cells were stimulated with Phytohaemagglutinin (PHA) (Sigma) of RPMI 1640 was added to each well. The mitogen PHA was added at final concentration of 5 μg/mL. Control wells contained 100 μL of PBMC suspension (1 × 106 viable cells/mL) without PHA (unstimulated). Cells were harvested at all times points of incubation period of 13 h.
PBMC cultures for HSP70.1 and HSP90.1 quantificationpresent.
Intracellular mRNAs for HSP70.1 and HSP 90.1 were evaluated in PBMCs (500 μL of cell suspension containing 1×106 cells/well) cultured under the conditions described above.
Analysis of HSP70.1 and HSP90.1 mRNA levels:
Levels of HSP70.1 and HSP90.1 mRNAs were measured by qRT PCR. Total RNA was isolated from unstimulated and stimulated cells of each heat treatments using Tri reagent (Sigma-Aldrich) following the procedure described by Singh et al. (2014) [30]. The purity and concentration of RNA was checked by a NanoDrop-2000 spectrophotometer (Thermo Fisher Scientific Inc. Wilmington, DE, USA). Prior to cDNA synthesis, RNA was treated with DNase treatment to remove any genomic DNA contamination. One μg of RNA was reverse transcribed using a RevertAid™ First Strand cDNA Synthesis Kit (M/s MBI Fermentas Life Sciences, Maryland, USA) according to the manufacturer’s protocol.
The resulting complimentary DNAs (cDNAs) were used in quantitative real-time polymerase chain reaction (qRT-PCR). The qRT-PCR for HSPs and housekeeping genes (Beta-actin) was performed in triplicate (technical replicate) using HotStart-IT SYBR Green qPCR Master Mix (2X) (Affymetrix), and primers for HSP70.1, HSP90.1 and Beta actin were used. Primers were used previously reported by Dangi et al. 2012 [31]. The sequences of primers are as follows: bovine Hsp70.1 5’-GACGACGGCATCTTCAAG-3’ (sense) and 5’-GTTCTGGCTGATGTCCTTC-3’ (antisense); HSP90.1- 5’-GCATTCTCAGTTCATTGGCTATCC-3’ (sense) 5’-GTCCTTCTTCTCTTCCTCCTCTTC-3’ (antisense) bovine Beta actin 5’- CTACCTTCAATTCCATCATG -3’(sense) and 5’- GCGATGATCTTGATCTTC-3’ (antisense) and Beta actin 5’- CTACCTTCAATTCCATCATG-3’ (Sense) and 5’-GCGATGATCTTGATCTTC. The predicted sizes of PCR products are 132 bp for HSP70.1; 190 bp for HSP90.1 and 163 bp for Beta actin. The cDNA samples were amplified in a real-time fluorescence thermal cycler (Fast 7500 Real Time PCR system, Applied Biosystem, USA) in 96 well plate. Thermal cycling conditions were 95 °C for 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 30 s. The relative expression of each sample was calculated using the 2−ΔΔCT method (Schmittgen and Livak 2008; Doley et al. 2014) [32,33]. Results were analyzed and shown as fold change (2−ΔΔCT) relative to the control group. All the reactions were performed in triplicate and the detected expression values were normalized against the endogenous control.
Statistical analysis:
All Real-time PCR reactions were run in triplicates. The statistical analysis was done using JMP 9.0 (JMP®, Version 9.0. SAS Institute Inc. Cary, NC, 1989–2007). As all the comparisons made above are pairwise comparisons, t-test was done to test the statistical significance.
Results and Discussion
In the present study, PBMCs isolated from Indian Tharparkar and Crossbred cows (Vrindavani) were utilized as cellular in-vitro model to assess the comparative cellular tolerance during heat stress. The transcriptional induction of HSP70 and HSP90 mRNAs was used as indicators to evaluate the comparative cellular tolerance ability of both the cattle types.
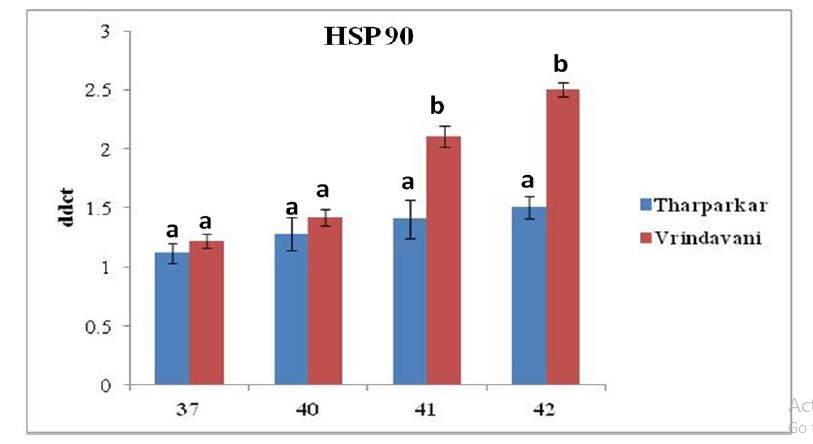
PMBCs were challenged with different high temperature at 37°C, 40°C, 41°C and 42°C for 65 h, respectively in two bovine species. The relative expression of HSP70.1 and HSP 90.1 increased significantly (P < 0.05) with increase in exposure temperature in both the cattle during the entire period of heat stress. Real-time PCR analysis revealed that the expression of HSP70.1 was upregulated by 1.00±0.10 at 37°C; 1.75±0.13 at 40°C ; 2.67±0.17 at 41°C and 3.08± 0.02 at 42°C and the expression of HSP90.1 by 1.11±0.08 at 37°C; 1.28±0.13; 1.40±0.16 at 41°C and 1.50± 0.09 at 42°C in Tharparkar cattle while in Vrindavani cattle, expression of HSP70.1 was up-regulated by 1.02±0.05 at 37°C; 2.74±0.07 at 40°C; 2.98±0.15 at 41°C and 3.25±0.13 at 42°C and HSP90.1 by 1.21±0.06 at 37°C; 1.41± 0.06 at 40°C; 2.11±0.09 at 41°C and 2.51±0.06 at 42°C. Increasing incubation temperature in both breeds increased the expression of both the HSPs genes, however, the expression of HSP70.1 and HSP90.1 were more pronounced in the PBMCs isolated from Vrindavani cattle than those from Tharparkar cattle (Figure 1).
High environmentally friendly temperatures directly or indirectly affects the efficiency of dairy cattle. Similarly, heavy beef cattle are unable to resist thermal stress compared to lighter cattle because high fat unseating averts heavy cattle from adaptable their body temperature proficiently (Pragna, et al. 2017).
A number of previous studies performed in bovine or other species estimated mRNA levels of HSP genes following exposure to heat stress. In our study, we have found that though the expression levels of both the HSP mRNAs were up-regulated in response to heat stress, it was more pronounced in PMBCs isolated from crossbred Vrindavani cow. PBMCs from Tharparkar cow expressed lower amount of both the HSPs at all temperatures. This pattern of heat stress response suggests that Indian native cattle breed Tharparkar is more thermotolerant than the crossbred cattle Vrindavani. Lower amount of HSP may results from lower protein denaturation and thus, better resistance to heat stress. Previous studies have reported the similar findings. A lower amount of HSP70 was detected in the two thermotolerant breeds (Brahman and Senepol cattle) when the isolated PMBCs were subjected to heat stress (Kamwanja et al. 1994; Mehla et al. 2014) [34,35]. Another study investigated it further and reported that reduced HSP70 expression in heat stressed Brahman and Senepol cattle might be indicative of reduced protein denaturation as denatured proteins serve as signal for HSP70 synthesis. Lacetera et al. (2006) [29] also observed that when PBMCs isolated from the Holstein cows given a chronic heat shock, produced less amount of HSP72 than those isolated from the Brown Swiss cows and predicted the lower tolerance of Brown Swiss cows was associated with higher expression of HSP72.
The expression profile of different HSP transcripts was measured in PBMCs during heat shock condition. The reason for selecting four major HSP genes (HSP70 and HSP90) for the present analysis was due to their primary role as molecular chaperons that ensures the correct protein folding and apoptosis regulation during physiological stressful conditions. It is also important that though heat shock response is an evolutionary conserved mechanism, different breeds/species may vary in their inherent ability to induce HSP synthesis during heat shock
However, these are several reports where high expression of HSP was co-related with increased thermotolerance (Angelidis et al. 1991; Horowitz 2002; Hansen 1999; Horowitz 2002; Meza-Herrera et al. 2006; Maibam et al. 2017) [36-39]. It has been observed that animals well adapted to warm climates produce lesser amounts of HSP proteins (Kamwanja et al. 1994; Lacetera et al. 2006; Romero et al. 2013) [34,29,40]. The activation of HSP genes is also related to the defense against cell damage from heat shock (Schiaffonati and Tiberio 1997; Rajhans et al. 2010) [41-48] and up-regulation of HSP72 expression might function as an indicator of cellular stress levels (Kristensen et al. 2004) [23].
Conclusion
In conclusion, our study found that bovine PBMCs in response to heat stress produced HSPs, but the expression levels of HSPs were different in both the cattle breeds and showed difference in their ability to ameliorate the deleterious effect of heat stress so as to maintain cellular integrity. PBMCs isolated from crossbred cattle produced higher expression levels of HSP70 as well as HSP90 than those isolated from heat-adapted Tharparkar cattle. Although heat shock response is an evolutionary conserved mechanism, different breeds/species as seen in the present study may vary in their inherent ability to induce HSP synthesis during heat shock. We suggest that increased thermotolerance of Tharparkar cattle is associated with the reduced production of HSPs, as expression of these stress proteins reflects extent of cellular damage.
Acknowledgments
The authors are grateful to the NICRA project, Indian Council of Agricultural Research, New Delhi, Government of India for financial assistance. I am highly thanks to ICAR-Indian Veterinary Research Institute (IVRI), Izatnagar, Bareilly, U. P.
- BAHS-Basic Animal Husbandry Statistics (2012) Department of Animal Husbandry, Dairying & Fisheries, Ministry of Agriculture, Government of India. Krishi Bhavan, New Delhi.
- Rajoriya JS, Prasad JK, Ghosh SK, Perumal P, Kumar Anuj Kaushal, Shobhana, Ramteke SS (2014) Studies on effect of different seasons on expression of HSP70 and HSP90 gene in sperm of Tharparkar bull semen. Asi Paci J Repro 3: 192.
- Sharma R, Maitra A, Singh PK, Tantia MS (2013) Genetic diversity and relationship of cattle populations of East India: distinguishing lesser known cattle populations and established breeds based on STR markers. SpringerPlus 2: 359.
- Sengar GS, Deb R, Singh U, Raja TV, Kant R, et al. (2018). Differential expression of microRNAs associated with thermal stress in Frieswal (Bos taurus x Bos indicus) crossbred dairy cattle. Cell Str. Chaper 23: 155-70.
- Hansen PJ (2004) Physiological and cellular adaptations of zebu cattle to thermal stress. Anim. Reprod Sci 82: 349-60.
- O’Bannon EB, Cornelison PR, Ragsdale AC, Brody S (1955) Relative growth rates at 80 and 50 F. of Santa Gertrudis, Brahman and Shorthorn heifers. J Anim Sci 14: 1187.
- Johnson HD (1965) Environmental temperature and lactation (with special reference to cattle). Int. J. Biometeor. 9: 103–16.
- Johnston JE, Naelapaa H, Frye JB (1963) Physiological responses of Holstein. Brown Swiss and Red Sindhi crossbreed bulls exposed to high temperatures and humidities. J Anim Sci 22: 432-36.
- Chen F, Qiu H (2012) Identification and characterization of a Chinese isolate of Cryptosporidium sepentis from dairy cattle. Parasitol Res 111: 1785-91.
- Singh AK, Upadhyay RC, Malakar D, Kumar S, Singh SV (2014) Effect of thermal stress on HSP70 expression in derimal fibroblast of zebu (Tharparkar) and crossbred (Karan-Fries) cattle. J Therm Bio 43: 46-53.
- Deb R, Sajjanar B, Singh U, Kumar S, Brahmane MP, et al. (2013) Promoter variants at AP2 box region of Hsp70.1 affect thermal stress response and milk production traits in Frieswal cross bred cattle. Gene 532: 230-5.
- Narayan U, Singh DV, Sharma RJ (2007) Physiological responses in Holstein–Friesian crossbred lactating cows under heat stress ameliorating treatments. Ind J Anim Res 41: 130–3.
- Deb R, Sajjanar B, Singh U, Kumar S, Singh R, et al. (2014) Effect of heat stress on the expression profile of Hsp90 among Sahiwal (Bos indicus) and Frieswal (Bos indicus* Bos taurus) breed of cattle: A comparative study. Gene 536: 435-40.
- Driver AM, Khatib H (2013) Physiology and Endocrinology Symposium: heat shock proteins: potentially powerful markers for preimplantation embryonic development and fertility in livestock species. J. Anim. Sci. 91: 1154-61.
- Mathew A, Morimoto RI (1998) Role of the heat‐shock response in the life and death of proteins. Ann. New York Aca Sci 851: 99-111.
- Li Z, Srivastava P (2004) Heat‐Shock Proteins. Current Protocols in Immunology, A-1T.
- Sodhi M, Mukesh M, Kishore A, Mishra BP, Kataria RS, et al. (2013) Novel polymorphisms in UTR and coding region of inducible heat shock protein 70.1 gene in tropically adapted Indian zebu cattle (Bos indicus) and riverine buffalo (Bubalus bubalis). Gene 527: 606-15.
- West JW (2003) Effects of heat stress on production in Dairy Cattle. J Dai Sci 86: 2131- 2144.
- Wheelock JB, Rhoads RP, VanBaale MJ, Sanders SR, Baumgard LH (2010) Effects of heat stress on energetic metabolism in lactating Holstein cows. J Dai Sci 93: 644–55.
- Hu H, Zhang Y, Zheng N, Cheng J, Wang J (2016) The effect of heat stress on gene expression and synthesis of heat-shock and milk proteins in bovine mammary epithelial cells. Anim Sci J 87: 84-91.
- Gill JK, Arora JS, Sunil Kumar BV, Mukhopadhyay CS, Kaur S, Kashyap N (2017) Cellular thermotolerance is independent of HSF 1 expression in zebu and crossbred non-lactating cattle. Int J Biometeor 61: 1687-1693.
- Bianca W (1961) Heat tolerance in cattle-its concept, measurement and dependence on modifying factors. Int. J. Biometeor 5: 5–30.
- Kristensen TN, Lovendahl P, Berg P, Loeschcke V (2004) HSP72 is present in plasma from Holstein-Friesian dairy cattle, and the concentration level is repeatable across days and age classes. Cell Str. Chaper 9: 143-9.
- Kumar A, Ashraf S, Goud TS, Grewal A, Singh SV, et al. (2015) Expression profiling of major heat shock protein genes during different seasons in cattle (Bos indicus) and buffalo (Bubalus bubalis) under tropical climatic condition. J. Therm. Bio. 51: 55-64.
- Verma N, Gupta ID, Verma A, Kumar R, Das R, et. Al. (2016) Novel SNPs in HSPB8 gene and their association with heat tolerance traits in Sahiwal indigenous cattle. Trop. Anim Heal Prod 48: 175-80.
- Kregel, K.C. (2002). Heat shock proteins: modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physio. 92: 2177-86.
- Picard D (2002) Heat-shock protein 90, a chaperone for folding and regulation. Cell. Mol Lif Sci 59: 1640-48.
- Singh RR, Dutt T, Kumar A, Tomar AKS, Singh M (2011) On-farm characterization of Vrindavani cattle in India. Ind J Anim Sci 81: 267–71.
- Lacetera N, Bernabucci U, Scalia D, Basirico L, Morera P, et al. (2006) Heat stress elicits different responses in peripheral blood mononuclear cells from Brown Swiss and Holstein cows. J Dai Sci 89: 4606–12.
- Singh LV, Saxena S, Gupta S, Gupta SK, Kumar G, et al. (2014) Evaluation and comparison of the constitutive expression levels of Toll-like receptors 2, 3 and 7 in the peripheral blood mononuclear cells of Tharparkar and Crossbred cattle from different regions of India. Vet Worl 7: 209-12.
- Dangi SS, Gupta M, Maurya D, Yadav VP, Panda RP, et al. (2012) Expression profile of HSP genes during different seasons in goats (Capra hircus). Trop Anim Heal Prod 44: 1905–12.
- Schmittgen TD, Livak KJ (2008) Analyzing real-time PCR data by the comparative C (T) method Nat Proto 3: 1101-8.
- Doley J, Singh LV, Kumar GR, Sahoo AP, Saxena L, et al. (2014). Canine parvovirus type 2a (CPV-2a)-induced apoptosis in MDCK involves both extrinsic and intrinsic pathways. Appl Biochem Biotech 172: 497-508.
- Kamwanja LA, Chase CC, Gutierrez JA, Guerriero V, Olson TA, et al. (1994) Responses of bovine lymphocytes to heat-shock as modified by breed and antioxidant status. J Anim Sci 72: 438-44.
- Mehla K, Magotra A, Choudhary J, Singh AK, Mohanty AK, et al. (2014) Genome-wide analysis of the heat stress response in Zebu (Sahiwal) cattle. Gene 533: 500-7.
- Angelidis CE, Lazaridis I, Pagoulatos GN (1991) Constitutive expression of heat shock protein 70 in mammalian cells confers thermo resistance. Eur J Biochem 199: 35-9.
- Horowitz M (2002) From molecular and cellular to integrative heat defense during exposure to chronic heat. Compar. Biochem. Physio 131: 475-83.
- Hansen PJ (1999) Possible roles for heat shock protein 70 and glutathione in protection of the mammalian preimplantation embryo from heat shock. Annu. Revi. Biomed Engin Sci 1: 5-29.
- Meza-Herrera CA, Martínez L, Aréchiga C, Bañuelos R, Rincon RM, et al. (2006) Circannual identification and quantification of constitutive heat shock proteins (HSP70) in goats. J. Appl Anim Res 29: 9–12.
- Romero RD, Pardo AM, Montaldo HH, Rodriguez AD, Hernández-Ceron J (2013) Differences in body temperature, cell viability, and HSP-70 concentrations between Pelibuey and Suffolk sheep under heat stress. Trop. Anim. Heal. Prod. 45: 1691-6.
- Schiaffonati L, Tiberio L (1997) Gene expression in liver after toxic injury: Analysis of heat shock response and oxidative stress inducible genes. Liver 17: 183-91.
- Rajhans R, Kumar GS, Dubey PK, Sharma GT (2010) Effect of timing of development on total cell number and expression profile of HSP-70.1 and GLUT-1 in buffalo (Bubalus bubalis) oocytes and pre-implantation embryos produced in vitro. Cell Biol. Inter 34: 463-8.
- Maibam U, Hooda OK, Sharma PS, Mohanty AK, Singh SV, Upadhyay RC (2017) Expression of HSP70 genes in skin of zebu (Tharparkar) and crossbred (Karan Fries) cattle during different seasons under tropical climatic conditions. J Therm Bio 63: 58-64.
- Doench JG, Petersen CP, Sharp PA (2003) siRNAs can function as miRNAs. Gen. Develop. 17: 438-42.
- Kumar R, Gupta ID, Verma A, Singh SV, Verma N, et al. (2016) Novel SNP identification in exon 3 of HSP90AA1 gene and their association with heat tolerance traits in Karan Fries (Bos taurus × Bos indicus) cows under tropical climatic condition. Trop. Anim. Heal. Prod. 48: 735-40.
- Leung AKL, Sharp PA (2010) MicroRNA functions in stress responses. Mol. Cell 40: 205–15.
- Li X, Cassidy JJ, Reinke CA, Fischboeck S, Carthew RW (2009) A microRNA imparts robustness against environmental fluctuation during development Cell 137: 273–82.
- Place RF, Noonan EJ (2014) Non-coding RNAs turn up the heat: an emerging layer of novel regulators in the mammalian heat shock response. Cell Str Chapero 19: 159-72.
FIGURE 1
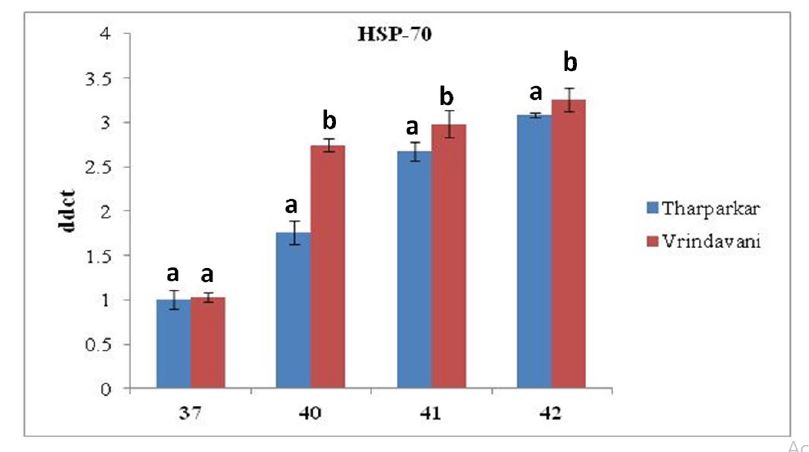
A: Assessment of relative mRNA expression level of HSP 70.1 gene in heat stressed PBMCs isolated from Tharparkar and Vrindavani cattle at 37°C, 40°C, 41°C and 42°C respectively. Data are represented as mean ± s.e.m. Levels (within time) not connected by same letter are significantly different (P<0.05).
B: Assessment of relative mRNA expression level of HSP 90.1 gene in heat stressed PBMCs isolated from Tharparkar and Vrindavani cattle at 37°C, 40°C, 41°C and 42°C respectively. Data are represented as mean ± s.e.m. Levels (within time) not connected by same letter are significantly different (P<0.05).
Figures at a glance