Comparative Transcriptomic Analysis Between SARS-COV-2, RSV And Influenza H3N2 Patients Highlights the Use Of IL-6 Inhibitors
Received Date: February 12, 2022 Accepted Date: February 14, 2022 Published Date: March 21, 2022
doi: 10.17303/jbcg.2022.5.102
Citation: Mondeali M, Bemani P, Hosseini P, Kesheh MM, Bahavar A et.al., (2022) Comparative Transcriptomic Analysis Between SARS-COV-2, RSV And Influenza H3N2 Patients Highlights The Use Of IL-6 Inhibitors. J Bioinfo Comp Genom 5:1-15
Abstract
Comparative transcriptomic analysis between different respiratory viral infections could help identifying the fundamental targets to improve the administration of these diseases. Especially in the case of COVID-19, unprecedented epidemy has challenged the health system with limited treatment options. Recently, based on studies on thousands of patients, WHO has recommended the use of IL-6 inhibitors in COVID-19. Here, we subjected the patient transcriptome of SARS COV2, RSV, and Influenza A to a comparative analysis. Two gene expression profiles were obtained from the NCBI GEO database. The differential analysis was performed by the R’s LIMMA package and Galaxy limma voom tool. To compare gene expression, Ggplot2 and pheatmap packages of R were applied to generate volcano plots and heatmap. Afterward, the KEGG pathway and GO enrichment were analyzed by DAVID online database. We used the STRING database for PPI network construction and hub gene identification. Using the DGIdb, potential drugs were predicted. We found a group of 11 genes, including IL6, IRF7, MX1, IFIH1, OAS1-3, XAF1, IRF9, IFIT1, and IFIT3, … and… with potentially the highest score in terms of gene expression level in all three viruses. IL-6 is particularly of higher importance with three FDA approved drugs, Siltuximab, Olokizumab, and Clazakizumab. Five other genes including, , IRF7, IFIH1, OAS1, IFIT3, and OAS3, also had potential therapeutic targets for SARS-CoV-2 and IAV H3N2 and RSV.
Keywords: SARS-CoV-2, IAV, RSV, IL6
Introduction
Viral respiratory infections have always been affected human’s life and being a challenge for public health. Only in around 100 years, we have encountered many epidemics of respiratory viruses such as different types ofInfluenza A (IAV), including; H1N1 in 1918, 2009 and 1977, H2N2 in 1957 and H3N2 in 1968. Another such epidemies are originated from coronal viruses, including SARS-CoV in 2002, MERS-CoV in 2012, and finally SARS-CoV-2 in 2019 [1-3]. Moreover, Respiratory Syncytial Virus (RSV) is another important virus, especially in young children aged under two years, elderly, and immunodeficient. RSV causes infections in the lower respiratory tract, which leads to bronchitis and pneumonia, although it can infect healthy adults asymptomatically [4].
After cell entry of respiratory viruses, secretion of IFNβ by the infected cells results in paracrine type I IFN signaling through the IFNα/β receptor, which induces hundreds of interferon-stimulated genes (ISGs) and inflammatory responses that have a crucial role for infection clearance. Still, a disturbance in immunity responses can be turned into a threat to the body. Cytokine storm induction is a common feature among these three viruses. These viruses can trigger the overexpression of inflammatory cytokines interleukin IL-1β, IL-6, IL-8, IL-12, Tumor Necrosis Factor (TNF-α), etc., from overactivated inflammatory cells such as neutrophils and macrophages, leading to the release of active mediators. Furthermore, effector cells such as T lymphocytes and NK cells are increased by the secreted cytokines to participate in tissue damage [5, 6].
Recent meta-analysis based on thousands of COVID-19 patients show the significance of IL-6 inhibition strategies in the treatment of this disease (ref: doi: 10.1001 / jama.2021.11330).
WHO has accordingly recommended IL-6 inhibitors in the management of OCVID-19 (https://www.who.int/news/item/06-07-2021-who-recommends-life-saving-interleukin-6-receptor-blockers-for-covid-19-and-urges-producers-to-join-efforts-to-rapidly-increase-access) In the current study we found interesting link between the transcriptome of SARS-CoV-2, RSV, and IAV H3N2. Especially, in the case of Covid-19 pandemic, it is of particular importance to study this disease in the context of other respiratory infections and learn from the experience of the decades of combat with these diseases and find proper possibilities in our currently approved antiviral arsenal. The comparative differentially expressed genes (DEGs) showed overlapping hub genes among three viruses for protein-protein interaction, and gene-drug interaction for available chemotherapeutic drugs to suggest potential drugs affecting these three viruses.
Materials
Data collection
Data analysis procedures are showed in Figure 1. Two gene expression profiles were obtained from the NCBI GEO database (https://www.ncbi.nlm.nih.gov/geo/): GSE32138 (GSM796520 to GSM796535) and GSE147507 (GSM4432378 to GSM4432383). The data for GSE147507 was generated by high throughput sequencing, Illumina NextSeq 500 method, while GSE32138 dataset was from microarray method using Agilent G2505C Microarray Scanner. GSE32138 contains triplicate primary human lung epithelium for SARS-CoV-2, and GSE147507 includes human airway epithelial cells for RSV and IAV H3N2 with a total of 3/3, 4/4, and 4/4 mock controls/infected cells culture, respectively, used for the generation of data in the interest datasets for DEGs analysis.
Sample Sources of the SARS-CoV-2, RSV, and IAV H3N2 Datasets
Generally, the primary target cells of the three viruses were respiratory cells. Triplicate primary human lung epithelium for SARS-CoV-2 and quadruplicate human airway epithelial cells for RSV or IAV H3N2 with 3/3, 4/4, and 4/4 mock controls/infected cells culture, respectively, were used for the generation of data in the datasets of interest. We selected all sixteen samples, GSM796520 to GSM796535, from the GSE32138 dataset and only six samples, GSM4432378 to GSM4432383, from the GSE147507 dataset for DEGs analysis.
DEGs identification in the Interest Datasets
In selected samples, we applied two tools for DEGs identification in both datasets; the online Galaxy tool [7] for GSE147507 and R version 1.2.5019 software for GSE32138. The log₂ fold change (logFC) ≥0.58 or ≤-0.58 were considered for significantly changed DEGs.
Identification of DEGs
Firstly, for microarray data (GSE32138), downloaded the series Matrix file of GSE32138 from the GEO database. Data were then normalized using the normalized quantiles function from R package ‘LIMMA’ from the Bioconductor project. Afterward, the LIMMA package managed the differential analysis by entering related codes into R (version 3.4.4). Galaxy limma voom tool (https://usegalaxy.org/) used for RNA-seq data (GSE147507) to Identification of DEGs. Adj P-value was supposed to avoid the occurrence of false-positive results. Genes with log FC ≥0.58 or ≤-0.58 and adj P-value < 0.05 were taken as differentially expressed genes between infected cells and mocks. Ggplot2 and pheatmap packages of R were applied to generate volcano plot and heatmap, respectively, for the visualization of the identified DEGs.
Gene Ontology and GO enrichment analysis
The Database for Annotation, Visualization, and Integrated Discovery (DAVID 6.8; available online: http://david.ncifcrf.gov) is an online database that facilitates users’ performance biological analysis from data collection. KEGG pathway and GO enrichment used for analyses were conducted with DAVID. P < .01 to considered statistically significant.
PPI network construction and hub gene (Protein‐protein interaction)
PPI networks of common-DEGs were analyzed by the search tool for retrieving interacting genes (STRING database, V10.5; http://string-db.org/) that predicted protein functional associations and protein-protein interactions. Subsequently, Cytoscape software (V3.5.1; http://cytoscape.org/) was applied to visualize and analyze biological networks and node degrees after downloading analytic results of the STRING database with a confidence score > 0.4.
Drug-hub gene interaction
Based on the hub genes, drugs were selected that served as promising targets using the Drug-Gene Interaction Database (DGIdb; http://www.dgidb.org/search_interactions; version 3.0.2 – sha1 ec916b2). In the present study, the final drug list included just drugs that the FDA permitted. The online tool named Cytoscape was used to construct the interaction network between the potential drugs and the hub genes.
Results
We identified 1682 DEGs for IAV H3N2, 2581 for RSV, and 265 for SARS-CoV-2 with p-value < 0.05. Among 1682 and 2581 DEGs for IAV H3N2 and RSV, 1100 and 1196 genes were upregulated, while 583 and 1387 genes were downregulated. Similar pattern was observedfor SARS-CoV-2, in which, out of 265 DEGs, 177 genes were upregulated and 88 genes were downregulated. For identification and visualization of DEGs with statistical significance, drew a volcano plot for each virus (Figure 2 A). Also, Figure 2 B shows the heatmap of 50 top DEGs in each understudied virus. According to our analysis represented in a Venn diagram (Figure 2 C), 63 DEGs shared between these three viruses that 62 genes were overexpressed and only one gene decreased. We listed these common DEGs in more detail in Table 1. The sixty-three/63 DEGs enriched in biological pathways are related to the immune system, response to stimulus, multi-organism process, signaling pathways, biological regulation, locomotion, single-organism process, and reproduction (Figure 3). The common property of IAV H3N2, RSV, and SARS-CoV-2 is the cytokine storm phenomena that trigger inflammation and pathological damages, especially in the respiratory tract through overexpression of inflammatory, antiviral genes, and immune cell proliferation [6]. We found involved interferon signaling genes as prominent among 63 DEGs.
PPI Network and GDI
Protein-protein interactions (PPIs) was investigated as one of the possible ways for the altered transcriptomes to cause the disease. To predict the protein interactions which were derived from DEGs, we submitted all 63 DEGs to the STRING database [8], and the interaction between proteins was constructed with 63 nodes corresponding to the 63 common DEGs between three viruses, 316 edges, average node degree 10, and PPI enrichment p-value <1.0e-16 (Figure 3). We also analyzed the shared DEGs by using the Cytoscape tool to visualize further gene interactions [9], in which the genes ranked based on score. The IL-6 and IRF7 genes were identified to contain the highest score among increased DEGs. The last column of Table 1 shows the score of 63 common DEGs between three viruses using Cytoscape analysis, in which 11 DEGs were identified as hub genes with high scores.
We also investigated the interaction of hub genes with available drugs and potential drug ability to construct the gene-drug interaction network using DGIdb. IL6, IRF7, IFIH1, OAS1, IFIT3, OAS3 were identified as potential targets for RSV, H3N2, and SARS COV2 infection treatment. Among potential drugs, Siltuximab, Olokizumab, Clazakizumab drugs had a therapeutic effect on soluble IL6 through direct interaction with IL-6 protein and abrogating several genes involved IL-6 signaling DGIdb found (Figure 3).
Discussion
In all three viruses the number of upregulated genes are higher than downregulated genes which is associated with… (is this a known phenomenon in virus infection due to release of cytokines for example? – ref?)
The innate immune system provides the first line of protection against viral infections. Invasion and damage of the intrinsic barrier on the airway epithelium, followed by respiratory viral infections such as SARS-CoV-2, IAV, and RSV, lead to the induction of several classes of PRRs, as the viral sensing pathways. Subsequently PRRs trigger, intracellular signaling cascades, and finally, activation of downstream antiviral responses, including the production of IFNs, ISG, and proinflammatory cytokines [10-12].
Rapid advances in microarray technologies and studies of host transcriptome would give scientists new and more comprehensive insights into the procedure to virus-host interactions that enable proposing potential/new therapeutic targets [13, 14]. In the current study, based on the updates in the GSEs (147507, 32138) dataset, our results show that 11 genes of the 63 common genes, including IL6, IRF7, MX1, IFIH1, OAS1-3, XAF1, IRF9, IFIT1, and IFIT3 had the highest score in term of gene expression level (Table 1). Moreover, according to the analysis listed in Table 1, the study revealed that these genes were upregulated in all three viruses.
IFN-I induces immune responses through JAK1/ TYK 2 activation followed by STAT1, STAT2, and IRF9. Also, IFN-1 is capable of activating (STAT3, 4, 5, and 6). It has been seen that when STAT1/2 and IRF9 transcription factors are activated, it leads to the induction of hundreds of IFN-stimulatory genes (ISGs) [15]. Our analysis showed that IRF9 was elevated during infection with the three viruses. We also detected an enhancement in the (IRF7) gene expression SARS-CoV-2 infection and two other respiratory viruses, RSV and Influenza A.
IRF7 plays a significant role in the expression of IFN-I [16]. Viral PAMPs are detected by PRRs such as (TLR3, TLR7/8) as well as the RIG-I-like receptor families (RIG-I and MDA5). This is followed by a cascade of the signaling pathway that leads to activation of downstream transcription factors, including IRF3, IRF7, and NF-κB, their translocation into the nucleus, and eventually transcription of IFN-I genes [17]. IFI16 and TLR2-mediated signaling pathways are also involved in activating IRF7 [18].
Upregulation of the IRF7 gene in response to RSV infection has been supported by the study conducted by McDonald et al. [19]. Similarly, Lowen and colleagues have shown that IRF7 was upregulated in response to IAV infection in guinea pigs as a host model of this virus [20].
The IFN type I produced by virus-infected cells in response to viral sensors such as intracellular TLRs, RIG-I, and MDA5 confers antiviral immunity in uninfected cells. For instance, 2’–5’-oligoadenylate synthetases (OAS1, OAS2, OAS3), which are the members of the OAS family, are induced by type I interferons in uninfected cells. They bind to viral dsRNA afterward activates RNase L, which degrades viral and cellular RNA and thereby limits viral replication [21, 22]. Consistent with other studies, our analysis showed increased levels of OAS1-3. For instance, it has been demonstrated that OAS1 was remarkably upregulated in human primary epithelial cells infected by IAV (H3N2) [23]. The overexpression of OAS1 in SARS-CoV2 and RSV infections has also been confirmed by Lieberman [24] and Ampuero investigations [25]. Moreover, several studies showed OAS2 was significantly upregulated in the acute phase of IAV, RSV, and SARS-CoV-2 infection [26-28]. In addition, up-regulation of the OAS3 gene was supported by McDonald’s study [19], and also Fjaerli et al. observation that represented the OAS3 gene up-regulation in the infected infants [29].
Another mechanism by which IFN-I confer antiviral IFIT genes in the un-infected cells. IFIT genes are distinguished and well-known ISGs with four family members, recognized in humans: IFIT1, IFIT2, IFIT3, and IFIT5 [30]. IFIT proteins are involved in antiviral activities mainly by different mechanisms, including restricting viral RNA translation, recognizing and sequestering the viral RNA, binding to the viral proteins, and reducing virus replication [31]. In the absence of stimuli, most cell types have silent or have a very low level of IFIT genes expression. In contrast, they are prominently induced by type I and type III interferons as strong and weak inducers, respectively. IFIT1 is also involved in inhibiting mRNA translation initiation through direct interactions with eIF3 and hence suppresses cells and virus’s protein synthesis. IFIT2 and IFIT3 are essential for modulate apoptosis and cell proliferation [32, 33]. Some observations close to our finding represented an increased expression of IFIT1 and 3 in virus-infected cells and patients [19, 25, 34-37].IFIH1, also known as MDA-5, is the cytoplasmic sensor that binds to the ds-RNA and, as mentioned above, as a part of innate immune response to viral infection triggers IFN-I induction. During the detection of double-stranded RNAs by the CTD and helicase domains of IFIH1, then conformational changes formed a filament of the molecules and association MAVS, leading to further signaling events that finally boost transcription of IFN-I and ISGs [38]. Our results show up-regulation at the IFIH1 level in agreement with Loganathan et al. study on SARS-CoV-2 infection [22], Cao et al., IAV (H3N2) infected cells, [39], and Okabayashi et al. in RSV infection [40].
The interferon-inducible MX1 gene codes a GTPase protein as part of the antiviral response induced by type I in the infected cells. Mx GTPases have antiviral activity against a large variety of RNA viruses, especially influenza viruses at the entry stage and before viral replication. Generally, Mx protein appears to recognize viruses by sensing nucleocapsid and subsequently captures viral ingredients in the host cells [41, 42]. Other studies confirm the elevated level of MX1 in our findings. For example, Barral-Arca revealed a significant increase in the MX1 gene expression levels when RSV infected patients were compared with healthy controls [43]. Consistently, the upregulation of the MX1 gene has been shown in the cells infected with various influenza subtypes A virus [44] and SARS-CoV-2 infection [22].
X-linked inhibitor of apoptosis XAF1is an IFN-induced gene that enhances IFN-mediated apoptosis and robustly affects IFN-induced sensitization of the cell to apoptosis. Studies have shown that XAF1 plays a role in pro-apoptotic responses. XAF1 is widely expressed in normal tissues at low basal levels can be upregulated in response to IFN and other cytokines like TNF-α. Early studies have revealed that XAF1 binding to XIAP inhibited XIAP’s anti-apoptotic activity [45, 46]. Our analysis close to Loganathan et al. study evaluating revealed an increase in the expression of XAF1 in SARS-CoV-2 infection [22]. IL-6 is a proinflammatory and pleiotropic cytokine produced by several cell types in response to cell damage, and infections control the expansion of chronic inflammatory diseases [47, 48]. For IL-6 signaling onset, first IL6 binds to the non-signal transducing IL-6R, followed by complex formation through gp130.
After IL-6 receptor complex formation, IL-6 initiates a cascade of signaling events primarily related to the JAK/STAT3 activation pathway [49]. Several studies have indicated a strong relationship between IL-6 levels in the serum and future respiratory failure [50]. Recent studies have shown the detrimental effects of IL-6 in viral clearance, supporting viral persistence and hence chronic infections via several mechanisms [51]. The results of our study showed an upregulation in IL6. The elevated level of this proinflammatory cytokine was in agreement with various studies on RSV [52], SARS-CoV-2 [53], and influenza [54] viral infections. During viral infections, IFN-I and inflammatory cytokines are produced by different cells to limit viral replication and its spreading. If this process is done well, the virus may be eliminated by the immune system. However, these antiviral processes may not turn off in a sensible time frame, do not occur at all, or function efficiently, in which case we will see dysfunction or tissue damage .]55[ One of the most important events during infection with respiratory viruses such as Influenza, SARS-COV-2, and RSV is cytokine storm, which causes damage to the lungs. During infection with these three viruses and the occurrence of cytokine storm phenomenon, various cytokines and chemokines are increased and lead to tissue damage.However, among the genes examined in the present study, IL6 is commonly elevated during infection with these viruses [56-58]. Since in the early stages of viral infection, the role of IFN and ISGs against viral infections and the future outcome seems very important, so we assume that they can be used potentially as diagnostic or prognostic markers or as targets for therapy for viruses such as Influenza A, SARS-COV-2, and RSV. Some of the 11 genes studied in this study, such as IL6, which plays a significant role in inflammation, severe tissue damage, and viral persistence, could be potentially used as targeted by appropriate therapeutic drugs. In the current study, Siltuximab, Olokizumab, and Clazakizumab drugs have been detected as an inhibitor of soluble IL6. Siltuximab is an FDA-approved chimeric human-mouse monoclonal antibody against IL-6 that forms high affinity and steady complexes with human IL-6. This drug prevents the binding of human IL-6 to both soluble and membrane-bound IL-6 receptors that can inhibit the formation of the hexameric signaling complex with gp130 on the cell surface and interdiction activating of the JAK-STAT signaling pathway [59]. Given that IL6 initiates the cytokine storm (58) and is a common gene with high expression level among the RSV, Influenza H3N2 and SARS COV2 preventing its production and signaling pathway could be considered for treatment of the patients infected with these three viruses.
Conclusion:
Our analysis identified 61 common DEGs between influenza A H3N2, RSV, and SARS Cov2.. Among them, eleven common hub genes might be the core genes, including IL6, IRF7, MX1, IFIH1, OAS1-3, XAF1, IRF9, IFIT1, and IFIT3. All of them were upregulated, and the overexpression of these genes was associated with inflammatory responses and cytokine storm that can lead to tissues damage. More importantly, IL6, IRF7, IFIH1,OAS1, IFIT3, OAS3 were identified as potential therapeutic targets. IL-6 with 3 FDA-approved drugs, Siltuximab, Olokizumab, and Clazakizumab could be considered as a high-scored potential target for upcoming COVID-19 clinical trials.
Acknowledgments
Not applicable.
Authors’ contributions
MM, PB and KHB designed the manuscript. MM, PB, PH, MMK, AB, AZ, TL, KHB, and AH wrote the first draft. PB and KHB edited the following versions. All authors read and approved the final manuscript.
Funding
The authors declare that no funding was received.
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
All authors approved the manuscript for publication.
Competing interests
The authors declare that they have no competing interests.
- Kilbourne ED, (2006) Influenza pandemics of the 20th century. Emerging infectious diseases. 12:9-14.
- Jain S, et.al., (2009) Hospitalized patients with 2009 H1N1 influenza in the United States, April–June 2009. New England journal of medicine. 361:1935-44.
- Rabaan AA, et.al., (2020) SARS-CoV-2, SARS-CoV, and MERS-COV: a comparative overview. Infez Med. 28: 174-84.
- Baker RE, et.al., (2019) Epidemic dynamics of respiratory syncytial virus in current and future climates. Nature Communications. 10:5512.
- Ragab D, et al., (2020) The COVID-19 Cytokine Storm; What We Know So Far. Frontiers in Immunology. 11(1446).
- Liu Q, Zhou YH, and Yang ZQ, (2016) The cytokine storm of severe influenza and development of immunomodulatory therapy. Cellular & Molecular Immunology. 13:3-10.
- Afgan E, et al., (2018) The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic acids research. 46:537-44.
- Szklarczyk D, et al., (2019) STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic acids research. 47:607-13.
- Shannon P, et al., (2003) Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome research,. 13:2498-04.
- Koyama S, et al., (2008) Innate immune response to viral infection. Cytokine. 43:336-41.
- Moriyama M, Hugentobler WJ and Iwasaki A, (2020) Seasonality of respiratory viral infections. Annual review of virology. 7:83-101.
- Troy NM and Bosco A, (2016) Respiratory viral infections and host responses; insights from genomics. Respiratory research. 17:1-12.
- de Lamballerie CN, et al., (2019) Characterization of cellular transcriptomic signatures induced by different respiratory viruses in human reconstituted airway epithelia. Scientific reports. 9:1-12.
- Tripp RA, Mejias A, and Ramilo O, (2013) Host gene expression and respiratory syncytial virus infection. Challenges and Opportunities for Respiratory Syncytial Virus Vaccines. 193-209.
- Teijaro JR, (2016) Type I interferons in viral control and immune regulation. Current opinion in virology. 16:31-40.
- Xue Q, et al., (2018) Seneca Valley Virus 3Cpro abrogates the IRF3-and IRF7-mediated innate immune response by degrading IRF3 and IRF7. Virology. 518:1-7.
- Yang S., et al., (2016) Interferon regulatory factor 3 is a key regulation factor for inducing the expression of SAMHD1 in antiviral innate immunity. Scientific reports. 6:1-16.
- Ning S, Pagano J and Barber GN, (2011) IRF7: activation, regulation, modification and function. Genes & Immunity.12:399-14.
- McDonald JU, et al., (2016) A simple screening approach to prioritize genes for functional analysis identifies a role for interferon regulatory factor 7 in the control of respiratory syncytial virus disease. Msystems. 1-3.
- Lowen AC, et al., (2007) Influenza virus transmission is dependent on relative humidity and temperature. PLoS Pathog. 3:151.
- Ibsen MS, et al., (2014) The 2’-5’-oligoadenylate synthetase 3 enzyme potently synthesizes the 2’-5’-oligoadenylates required for RNase L activation. Journal of virology. 88:14222-31.
- Loganathan T, et al., (2020) Host transcriptome-guided drug repurposing for COVID-19 treatment: a meta-analysis based approach. PeerJ. 8:9357.
- Zhou A, et al., Comprehensive Transcriptomic Analysis Identifies Cellular Immune Response Against Influenza A Virus Infection. Available at SSRN 3736007.
- Lieberman NA, et al., (2020) In vivo antiviral host response to SARS-CoV-2 by viral load, sex, and age. BioRxiv.
- Ampuero S, et al., (2018) Time-course of transcriptome response to respiratory syncytial virus infection in lung epithelium cells. Acta virologica. 62:310-25.
- . Ioannidis I, et al., (2012) Plasticity and virus specificity of the airway epithelial cell immune response during respiratory virus infection. Journal of virology. 86:5422-36.
- Zhai Y, et al., (2015) Host transcriptional response to influenza and other acute respiratory viral infections–a prospective cohort study. PLoS Pathog. 11:1004869.
- Zhou S., et al., (2021) A Neanderthal OAS1 isoform protects individuals of European ancestry against COVID-19 susceptibility and severity. Nature medicine.1-9.
- Fjaerli HO, et al., (2006) Whole blood gene expression in infants with respiratory syncytial virus bronchiolitis. BMC infectious diseases 2006. 6:1-7.
- Pidugu VK, et al., (2019) Emerging functions of human IFIT proteins in cancer. Frontiers in molecular biosciences.6:48.
- Diamond MS and Farzan M (2013) The broad-spectrum antiviral functions of IFIT and IFITM proteins. Nature Reviews Immunology. 13:46-57.
- Fensterl V. and Sen GC, (2015)Interferon-induced Ifit proteins: their role in viral pathogenesis. Journal of virology 89:2462-68.
- Zhou X, et al., (2013) Interferon induced IFIT family genes in host antiviral defense. International journal of biological sciences. 9:200.
- Blanco MD, et al., (2020) Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 2020. 181:1036-45.
- Cao Y., et al., (2017) Differential responses of innate immunity triggered by different subtypes of influenza a viruses in human and avian hosts. BMC medical genomics. 10:41-54.
- Pennings JL., et al., (2018) Transcriptomics in lung tissue upon respiratory syncytial virus infection reveals aging as important modulator of immune activation and matrix maintenance. Scientific reports. 8:1-13.
- Rosa B.A, et al., (2021) IFN signaling and neutrophil degranulation transcriptional signatures are induced during SARS-CoV-2 infection. Communications biology. 4:1-14.
- Gorman JA, et al., (2017) The A946T variant of the RNA sensor IFIH1 mediates an interferon program that limits viral infection but increases the risk for autoimmunity. Nature immunology. 18:744.
- Cao Y, et al., (2020) Single-cell analysis of upper airway cells reveals host-viral dynamics in influenza infected adults. BioRxiv.
- Okabayashi T, et al., (2011) Type-III interferon, not type-I, is the predominant interferon induced by respiratory viruses in nasal epithelial cells. Virus research. 160:360-66.
- Haller O, Staeheli P and Kochs G, (2007) Interferon-induced Mx proteins in antiviral host defense. Biochimie. 89: 812-818.
- Verhelst J, et al., (2012) Interferon-inducible protein Mx1 inhibits influenza virus by interfering with functional viral ribonucleoprotein complex assembly. Journal of virology,. 86:13445-55.
- Arca RB, et al., (2020) A meta-analysis of multiple whole blood gene expression data unveils a diagnostic host-response transcript signature for respiratory syncytial virus. International journal of molecular sciences. 21:1831.
- Cao Y, et al., (2017) Differential responses of innate immunity triggered by different subtypes of influenza a viruses in human and avian hosts. BMC Med Genomics. 10:70.
- Lin B, Xu D, Leaman DW, (2016) X‐linked inhibitor of apoptosis‐associated factor 1 regulates TNF receptor 1 complex stability. FEBS letters. 590:4381-92.
- Jeong SI, et al., (2018) XAF1 forms a positive feedback loop with IRF-1 to drive apoptotic stress response and suppress tumorigenesis. Cell Death Dis. 9:806.
- Baran P, et al., (2018) The balance of interleukin (IL)-6, IL-6· soluble IL-6 receptor (sIL-6R), and IL-6· sIL-6R· sgp130 complexes allows simultaneous classic and trans-signaling. Journal of Biological Chemistry. 293:6762-75.
- Chiaretti A, et al., (2013) IL-1β and IL-6 upregulation in children with H1N1 influenza virus infection. Mediators of inflammation.
- Salinas VL, et al., (2019) The role of interleukin 6 during viral infections. Frontiers in microbiology. 10:1057.
- Gubernatorova E, et al., (2020) IL-6: relevance for immunopathology of SARS-CoV-2. Cytokine & growth factor reviews. 53:13-24.
- Salinas VL, et al., (2019) The Role of Interleukin 6 During Viral Infections. Front Microbiol. 10:1057
- Wiseman DJ, et al., (2020) Immunological and Inflammatory Biomarkers of Susceptibility and Severity in Adult Respiratory Syncytial Virus Infections. The Journal of infectious diseases. 222:S584-S591.
- Zhu L, et al., (2020) Single-cell sequencing of peripheral mononuclear cells reveals distinct immune response landscapes of COVID-19 and influenza patients. Immunity. 53:685-96.
- Matos ADR, et al., (2019) Antiviral potential of human IFN-α subtypes against influenza A H3N2 infection in human lung explants reveals subtype-specific activities. Emerging microbes & infections. 8:1763-76.
- Spencer JV, Religa P, Lehmann MH, (2020) Cytokine-Mediated Organ Dysfunction and Tissue Damage Induced by Viruses. Frontiers in immunology. 11:2.
- Hu B, Huang S, Yin L, (2021) The cytokine storm and COVID‐19. Journal of medical virology. 93:250-56.
- Bohmwald K, et al., (2019) Contribution of cytokines to tissue damage during human respiratory syncytial virus infection. Frontiers in immunology. 10:452.
- Xi-zhi JG, Thomas PG. (2017) New fronts emerge in the influenza cytokine storm. in Seminars in immunopathology. Springer.
- Pastor P T, Briz EL, Andrés JLP,( 2020) Involvement of interleukin 6 in SARS-CoV-2 infection: siltuximab as a therapeutic option against COVID-19. European Journal of Hospital Pharmacy. 27:297-98.
FIGURE 1
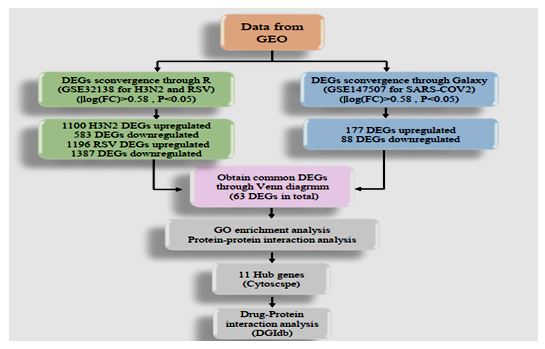
Figure 1: Flowchart of data preparation, processing, and analysis in this study
FIGURE 2
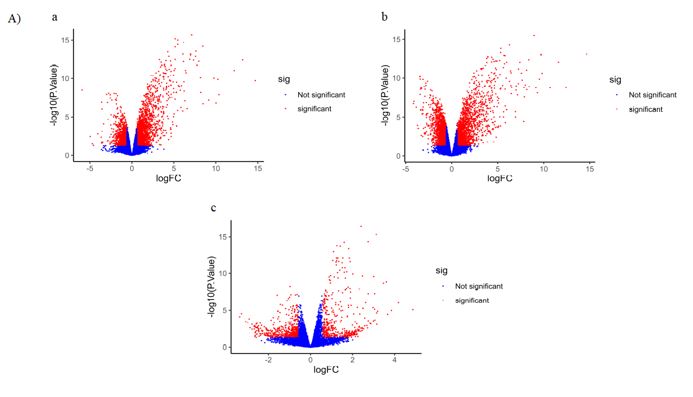
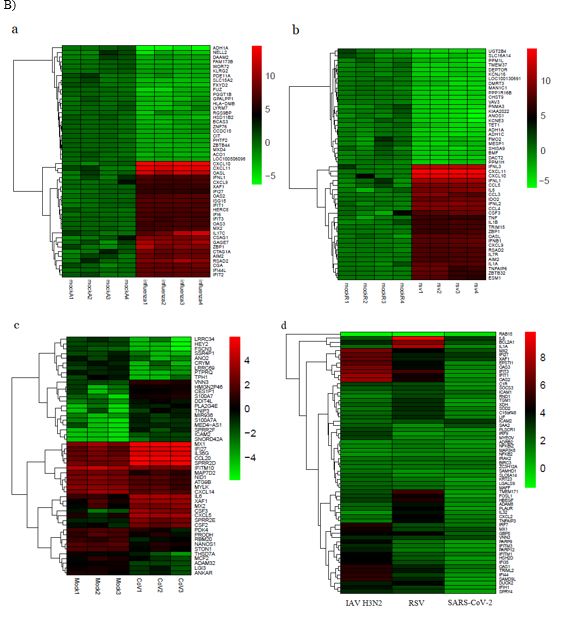
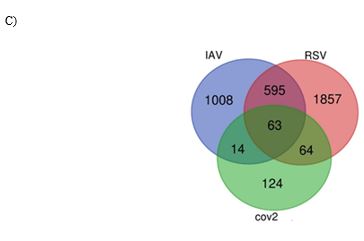
Figure 2: A) The volcano plots presenting DEGs for IAV H3N2 (a), RSV (b), and SARS-CoV-2 (c). The y-axis demonstrates the log10 values of the p-values for the significance of each examined DEG, while the x-axis shows the log10 values of fold changes for each DEG. Red plots depict DEGs with logFCs =0.58 or =-0.58, and blue plots displays remained DEGs with no statistical significance. B) Venn diagram of overlapping DEGs for three viruses. C) The heatmap of 50 top DEGs (25 increased and 25 decreased) in IVA H3N2 (a), RSV (b), SARS-CoV-2 (c), and 63 common DEGs (d). Red and green colors reflected upregulation and downregulation, respectively
FIGURE 3
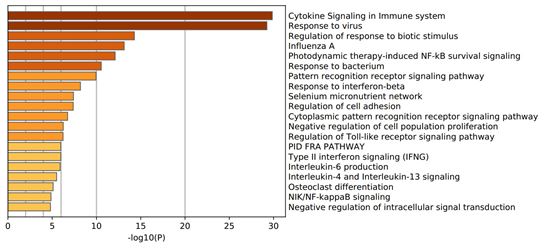
Figure 3: Gene Ontology (GO) enrichment analysis of the common differentially expressed genes
FIGURE 4
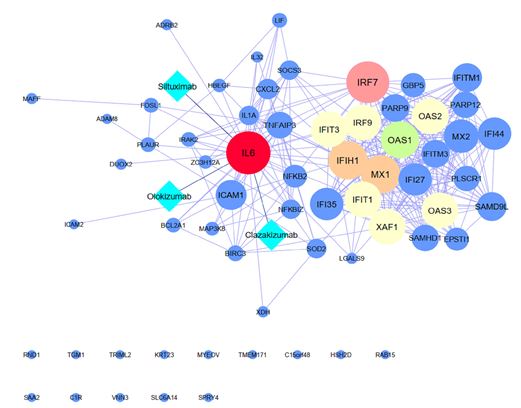
Figure 4: Protein-protein interactions network construction and Drug prediction with p-value <1.0e-16
Tables at a glance
Figures at a glance