Formononetin Inhibits the Development of NSCLC by Targeting HK1 to Reduce Glycolysis Levels
Received Date: April 10, 2024 Accepted Date: May 10, 2024 Published Date: May 13, 2024
doi: 10.17303/jcrto.2024.12.202
Citation: Pu Jiang, Xiao-peng Wang, Fu-hui Zhang, Sheng-jie Luan, Zi-rui Tan (2024) Formononetin Inhibits the Development of NSCLC by Targeting HK1 to Reduce Glycolysis Levels. J Cancer Res Therap Oncol 12: 1-13
Abstract
Background: This study aims to explore the mechanism of formononetin in preventing the occurrence of non-small cell lung cancer(NSCLC).
Methods: To explore the sequence of COX-2 overexpression and the occurrence of NSCLC through animal experiments, and verify the effect of formononetin in preventing the occurrence of NSCLC. To explore the mechanism of formononetin in NSCLC in cell experiments.
Results: The expression of COX-2 increased at 15 weeks (precancerous lung tissue) and 20 weeks (lung cancer tissue) after urethane treatment. The incidence of lung cancer and the expression of COX-2 in the tissues of mice treated with formononetin were significantly reduced compared with control. Formononetin inhibited the proliferation of NSCLC cells in a dose-dependent manner. Formononetin inhibited the cell cycle of NSCLC cells, cells in G1 phase increased significantly, while cells in S phase decreased significantly. Formononetin significantly inhibited glycolysis of NSCLC cells. Formononetin significantly reduced the expression levels of COX-2, cyclin D1 and HK1 in NSCLC cells and HK1 is a direct target of formononetin.
Conclusion: Formononetin has an anti-NSCLC effect by inhibiting aerobic glycolysis. It reversed the progression of NSCLC before the occurrence of NSCLC by suppressing inflammation.
Keywords: NSCLC; Formononetin; COX-2; glycolysis; HK1
Introduction
Lung cancer is the most common malignant tumor in the world, and it is also the main cause of death from malignant tumors in the world [1]. Lung cancer is mainly divided into non-small cell lung cancer (NSCLC) and small cell lung cancer (SCLC). Among them, non-small cell lung cancer accounts for 85% of lung cancers and its main pathological types are squamous cell carcinoma and adenocarcinoma [2]. Although the diagnosis and treatment of NSCLC have made significant progress, the 5-year survival rate is still only 15-20% [3,4]. Preventing the occurrence of NSCLC is the main way to reduce the incidence of NSCLC and relieve the pressure on the social medical system. Studying the pathogenesis of NSCLC helps to find ways to prevent NSCLC.
Some traditional Chinese medicine compound prescriptions have been proven to inhibit the progression of lung cancer. Lin [5] and Jiang’s [6] research showed that Yangzheng Xiaoji has a significant role in reducing the migration and invasion of lung cancer. The main ingredient of Yangzheng Xiaoji is astragalus, and formononetin, an isoflavone component isolated from astragalus, is reported to have an inhibitory effect on malignant tumors [7,8]. Formononetin may be the molecular basis for Yangzheng Xiaoji to exert its anti-cancer effect. Formononetin has various effects such as anti-inflammatory and anti-oxidative stress and studies have reported that formononetin could inhibit inflammation in mice by inhibiting COX-2 [9,10]. COX-2 is a common inflammatory index protein and an important ratelimiting enzyme in prostaglandin synthesis which can interfere with the angiogenesis and metastasis of malignant tumors [11,12]. Some studies have shown that inhibiting COX2 could inhibit the progression and metastasis of NSCLC [13,14]. Inflammatory factors are closely related to the pathogenesis of NSCLC [15]. However, the causal relationship between the overexpression of COX-2 and the occurrence of NSCLC is not clear. Whether the overexpression of COX-2 leads to the occurrence of cancer or the progress of cancer induces the overexpression of COX-2 remains to be studied. If the overexpression of COX-2 precedes the occurrence of NSCLC, can drugs inhibit COX-2 to prevent the occurrence of NSCLC? In this study, we verified the sequence between COX-2 overexpression and the occurrence of NSCLC in animal experiments, and verified that whether formononetin could prevent the occurrence of NSCLC by inhibiting COX-2 mediated inflammation.
Materials and Methods
Animals and Experimental Design
All experimental animals were C57BL/6 mice purchased from Beijing Vital River Laboratory Animal Technology. All mice were treated with urethane (Sigma, USA) in drinking water from 6 weeks of age to establish a lung cancer model. The concentration of urethane in drinking water is 20mg/100ml. The animals were weighed weekly and monitored daily. From each group, 12 mice were sacrificed at 15 and 20 weeks after chemical carcinogen treatment, and were sacrificed for tumor evaluation and related gene expression profiling studies.
Immunohistochemistry
The COX-2 expression levels were determined by IHC. The reagents were purchased from Shanghai Lanyi Technology Co., Ltd., and the kit instructions were strictly followed. Make paraffin sections, bake them for later use, dewax and hydrate with conventional xylene, block and inactivate endogenous peroxidase, then perform antigen retrieval, use citrate buffer as the antigen retrieval solution, and add 10% to the sections. The normal goat serum was blocked to reduce the background, the phosphate buffered saline (PBS) was removed after the serum was removed, followed by anti-COX-2 antibody (1:250; CST, USA) overnight at 4°C. After cleaning with PBS, the sections were then incubated with HRP-conjugated anti-rabbit IgG (ZSGB-BIO, China) for 45 min at 37°C. Then, 0.04% diaminobenzidine and 0.03% hydrogen peroxide solution were added for color development, and the staining depth was controlled under a microscope. Rinse with distilled water to terminate the staining, and counterstain with hematoxylin for 30 s. After rinsing with double distilled water for several times, it was dehydrated and transparent, and then observed under a microscope.
Cell Lines
A-549 and HCC827 cell lines were obtained from China Infrastructure of Cell Line Resources.
CCK-8 Assays
Pre-experimental results showed that when the concentration of Formononetin was 0.001uM/ml, there was no significant difference between the control groups in CCK-8 results, while the results showed significant difference when the concentration was 0.003uM/ml. The control cells were completely overgrown at 72 hours, so the experiment time was set to 48 hours. The cells were seeded in a 96-well plate at 2×107/L, 100 μL per well, and 5 replicate wells in each group. After 24 hours, remove the 96-well plate on which the cells have adhered, discard the old medium, and add different concentrations of Formononetin (0.003uM/ml and 0.006uM/ml) for 12, 24, 36and 48 hours respectively. The control group only contained cells and culture medium. Then add 100 μL of 10% CCK-8 working solution to each well. After completion, incubate in an incubator for 4 hours, and measure the absorbance (OD) value with a microplate reader at a wavelength of 450 nm.The experiment was repeated 3 times in parallel.
Flow Cytometry (FCM) Assays
After trypsinization to collect the pretreated cells, the cells were washed by PBS and resuspended to adjust the cell concentration to 1×109/L. Add pre-cooled 70% ethanol to 1×106 cells, fix overnight at 4℃, remove the fixative and wash the cells twice with pre-cooled PBS. Resuspend the cells with 100 μL RNase A solution. After mixing, incubate in a 37℃ water bath for 30 min. Add 400μLPI (Propidium Iodide), mix thoroughly and stain, and react for 30 min at 4℃ in the dark. Flow cytometer test on the machine, recording the red fluorescence at the excitation wavelength of 488nm
Seahorse Assays
After treatment with formononetin for 48 hours, A-549 and HCC827 cells were plated in XF96 Cell Culture Microplates at an initial cellular density of 1 × 104 cells/well the day before determination. A Seahorse Extracellular Flux (XF96e) Analyzer and the Agilent Seahorse XF Glycolytic Rate Assay Kit were used to measure the extracellular acidification rate (ECAR), reflecting the glycolytic level of live EC cells. The specific experimental procedures were performed according to the manufacturer's instructions.
Western Blot Analysis
After the pre-treated cells were digested and centrifuged, they were washed 3 times with pre-cooled PBS, mixed with 1× protein extract according to the number of cells, and incubated on ice for 30 min. Centrifuge at 12 000 × g for 10 min in a low-temperature centrifuge. After aspirating the supernatant and mixing it with the loading buffer, boiling water for 10 minutes. After preparing 8% (AcrBis) separation gel and concentrated gel, load the same amount, prepare fast electrophoresis solution, and turn on the power for electrophoresis, 140 V for 30 min. After the electrophoresis, the gel was cut and transferred, and the film was transferred at a constant current of 300 mA for 90 minutes. After the transfer, the film was sealed with 5% skimmed milk powder for 1 hour, and the cleaning solution (Tris-Buffered Saline 20, TBST) was washed 4 times on a shaker, 5 min each time. The membranes were incubated with antibody (anti-COX-2, 1:1,500, CST, USA; anti-CyclinD1, 1:1,500, CST, USA; anti-HK1, 1:1,000, CST, USA; anti-GAPDH, 1:5,000, SIGMA, USA) at 4°C overnight, washed three times with TBST. Incubate the polyvinylidene fluoride membrane (PVDF) with the diluent of the secondary antibody for 1 hour again, wash with the TBST cleaning solution 5 times, prepare the color mixing solution, incubate the PVDF membrane with the color solution for 30 s, and develop on the imager
Statistical Analysis
The experimental data was statistically analyzed with Excel and SPSS 21.0 software. The quantitative results are shown as mean±SD. The comparison of the rates between groups adopts the χ2 test or Fisher's exact probability method
Results
The Formononetin Treatment Reduced the Incidence of Lung Cancer and Decreased the Expression of COX-2 in Mice
There was no accidental death of mice in each group. The mice in the each group gradually gained weight and breathed normally. The weight difference began to appear in the 14th week after the start of cancer induction, and the formononetin group was significantly higher than the control group (Figure 1A).
As shown in the experimental, the histological examination revealed that lungl cancer was detected in control mice at 15 weeks (3/12), and was not detected in formononetin-treated mice at 15 weeks (0/12). And the cut-off date was at the 20th week after urethane-treated. The incidence of formononetin-treated mice was significantly lower than that of control mice at the 20th week (5/12 VS 11/12, P﹤0.05) (Figure 1B).
To investigate the levels of key inflammation markers during cancer development, IHC was performed. Both the precancerous lung (15 weeks) and the lung cancer tissues (20 weeks) showed high expression of COX-2 in control mice, COX-2 expression levels have increased before the occurrence of lung cancer. The expression of COX-2 in formononetin-treated mice was decreased compared with controls both in precancerous lung (15 weeks) (COX-2 positive rate: 2/12 VS 8/12, P﹤0.05) and the lung cancer tissues (20 weeks) (COX-2 positive rate: 6/12 VS 12/12, P﹤0.05) (Figure 1C-D).
The Effect of Formononetin on NSCLC Cells
To study the mechanism by which formononetin inhibits lung cancer progression, the NSCLC cell lines A-549 and HCC827 were treated with different concentrations of 0.003uM/ml and 0.006uM/ml formononetin, respectively, for 24,36,48 hours. Results showed that formononetin inhibited the proliferation of NSCLC cells in a dose-dependent manner. At 48 hours, 0.003uM/ml and 0.006uM/ml of formononetin reduced the viability by 45% and 56% in A-549 cell line, and by 36% and 53% in HCC827 cells (Figure 2A). When the concentration of formononetin is 0.003uM/ml, it can not only significantly inhibit the proliferation of NSCLC cell lines, but also maintain the activity of NSCLC cell lines, which is conducive to the follow-up experiment. Therefore, 0.003uM/ml is used as the experimental concentration in the follow-up experiment. NSCLC cell lines A-549 and HCC827 were treated with a concentration of 0.003uM/ml formononetin for 48 hours, respectively. According to the results of flow cytometry analysis, cells in G1 phase was increased by 11%, while cells in S and G2 phase was decreased significantly in A-549 cell line treated with formononetin. And in HCC827 cell line, cells in G1 phase was increased by 12% and cells in S and G2 phase was decreased significantly with formononetin treated (Figure 2B).
The Mechanism of Formononetin on NSCLC Cells
A-549 and HCC827 were treated with 0.003uM/ml formononetin. RT-qPCR was used to detect the expression levels of CDK4, CDK6, CyclinD1 and COX-2, the key genes in the cell cycle transition from G1 to S. The results showed that the expression levels of CyclinD1 and COX-2 mRNAs were decreased by formononetin and the levels of CDK4 and CDK6 were no change both in A-549 and HCC827 cells (Figure 3A). We further tested the expression levels of CyclinD1 and COX-2 proteins by Western Blot, and the results showed that the expression of CyclinD1 and COX-2 proteins were also decreased by formononetin both in A-549 and HCC827 cells (Figure 3B).
Formononetin Targets and Decreases the Expression of HK1
We searched the target genes of formononetin through TCMSP and PharmMapper and found that formononetin could target HK1 protein. HK1 is the rate-limiting enzyme of glycolysis. We simulated the interaction between formononetin and HK1 through molecular docking. Formononetin could be linked with HK1 through hydrogen bonds located at the 105th and the 253th THR (Figure 4A). The binding energy is -7.8 kcal/mol, which indicates that it could bind spontaneously and that the binding is stable. To determine how the HK1 protein was changed by formononetin treatment, we detected the expression level of HK1 protein, and the results showed that formononetin significantly inhibited the expression level of HK1 protein (Figure 4B). Then, we tested the effects of formononetin on the level of aerobic glycolysis by ECAR. The results showed that formononetin significantly inhibited aerobic glycolysis levels in both A-549 and HCC827 cells (Figure 4C).
Discussion
Chronic inflammation is closely related to the development of lung cancer [16-18]. COX-2 is an important protein associated with inflammation and tumors [19-21]. Jin et al showed that in the process of inducing the esophageal cancer in mice, the elevated level of COX-2 expression preceded the occurrence of cancer [22]. We first conducted a mice cancer induction experiment to detect the expression level of COX-2 in mice lung tissue at different times during the onset of lung cancer. In the experimental group, formononetin was added to mice to test its effect on preventing lung cancer. At 15 weeks (precancerous lung tissues) and 20 weeks (lung cancer tissues) after urethane-treatment, IHC showed that the expression of COX-2 in lung tissues or lung cancer tissues increased. The results show that: before the occurrence of lung cancer, the expression level of COX-2 has increased. This is consistent with the results of Jin et al. Compared with the control group, the incidence of lung cancer and the expression of COX-2 in the tissues of the mice treated with formononetin were significantly reduced. The experimental results show that formononetin can inhibit the occurrence of lung cancer by reducing the expression of COX-2.However, the mechanism by which formononetin acts on the pathogenesis of NSCLC through COX-2 is still unclear.
In order to explore the mechanism of formononetin on NSCLC, we further carried out cell experiments. First, we verified that formononetin has an inhibitory effect on NSCLC cell lines. We treated NSCLC cell lines A-549 and HCC827 with concentrations of 0.003uM/ml and 0.006uM/ml formononetin for 12, 24, 36, and 48 hours, respectively. According to the results of CCK-8, we found that formononetin inhibited the proliferation of NSCLC cells in a dose-dependent manner. COX-2 has been shown to promote the progression of lung cancer by promoting the cell cycle [23]. Therefore, we further examined the effect of formononetin on the cell cycle. NSCLC cell lines A-549 and HCC827 were treated with formononetin at a concentration of 0.003uM/ml for 48 hours. According to the results of flow cytometry analysis, compared with the control group, the cells treated with formononetin in the G1 phase increased significantly, while the cells in the S phase decreased significantly [24]. Cells enter the cell cycle indefinitely, leading to enhanced cell proliferation, which plays an important role in the occurrence of cancer. Studies have shown that arrest non-small cell lung cancer cells in G1 phase can significantly inhibit the proliferation of cancer cells [25-27]. Our experimental results show that formononetin can inhibit cancer cell proliferation by preventing cancer cells from entering the cell cycle. The results were consistent with our expectations that formononetin may inhibit the cell cycle by inhibiting COX-2 expression.
In order to explore how formononetin prevents cancer cells from entering the cell cycle. We used formononetin to process A-549 and HCC827.RT-qPCR and Western blotting were used to detect the expression levels of CDK4, CDK6, CyclinD1 and COX-2, the key genes in the cell cycle transition from G1 to S. The results showed that compared with the control group, formononetin significantly reduced the expression levels of COX-2 and Cyclin D1 mRNA and protein in the A-549 and HCC827 cell lines. Cyclin D1 is an important protein for cell cycle regulation, and it plays a central role in the cancer pathogenesis of immortal proliferation of cancer cells [28]. Uncontrolled transcription, accumulation and ubiquitination of Cyclin D1 and its homologous CDK can lead to uncontrolled cell growth. Therefore, Cyclin D1 is considered to be a carcinogenic driving factor for various cancers such as lung cancer, breast cancer and melanoma [29]. According to the results of animal and cell experiments, it was confirmed that formononetin can inhibit the expression of COX-2 and Cyclin D1 in NSCLC cells, thereby arresting the cells in the G1 phase, thereby inhibiting the occurrence of NSCLC.
We further analyzed the direct targets of formononetin and found that formononetin could bind to HK1 spontaneously and stably through hydrogen bonding and decrease the expression of HK1.HK1 is the rate-limiting enzyme of glycolysis, and formononetin inhibits glycolysis by inhibiting HK1. The G1 phase of the cell cycle is the early stage of DNA synthesis. The main function of G1 is to synthesize RNA and ribosomes, which require a large amount of energy and materials. Aerobic glycolysis can produce a large amount of ATP quickly, as well as a large number of metabolites, which can provide energy and materials for tumor progression [30]. These results explained the mechanism by which formononetin arrested the cell cycle in the G1 phase. Glycolysis is inhibited by formononetin, resulting in insufficient energy and material supply, what arrested the cell cycle in the G1 phase. At the same time, increased levels of glycolysis could enhance inflammation [31]. Formononetin could reduce inflammation by inhibiting glycolysis.
In short, it was demonstrated that formononetin has an anti-NSCLC effect by inhibiting aerobic glycolysis. It reversed the progression of NSCLC before the occurrence of NSCLC by suppressing inflammation, indicating that formononetin has the potential to be a drug that prevents NSCLC.
Data Availability Statement
The data utilized in this study are authentic and valid. The authors take full responsibility for their accuracy and reliability. They are available for public access upon request from the corresponding author.
Conflicts of Interest Statement
No conflict of interest exits in the submission of this manuscript, and manuscript is approved by all authors for publication.
Ethics Approval and Consent to Participate
The current study was approved by the Institutional Human Ethics Committee of Hebei Medical University Fourth Hospital (Shijiazhuang, China)
- Kratzer TB, Bandi P, Freedman ND, et al. (2024) Lung cancer statistics, 2023. Cancer, 130: 1330-48.
- Molina JR, Yang P, Cassivi SD, Schild SE and Adjei AA (2008) Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship. Mayo Clin Proc, 83: 584-94.
- Torre LA, Siegel RL, Ward EM and Jemal A (2016) Global Cancer Incidence and Mortality Rates and Trends--An Update. Cancer Epidemiol Biomarkers Prev, 25: 16-27.
- Travis WD, Brambilla E, Nicholson AG, et al. (2015) The 2015 World Health Organization Classification of Lung Tumors: Impact of Genetic, Clinical and Radiologic Advances Since the 2004 Classification. J Thorac Oncol, 10: 1243-60.
- Ye L, Ji K, Frewer N, Ji J and Jiang WG (2012) Impact of Yangzheng Xiaoji on the adhesion and migration of human cancer cells: the role of the AKT signalling pathway. Anticancer Res, 32: 2537-43.
- Jiang WG, Ye L, Ruge F, et al. (2015) YangZheng XiaoJi exerts anti-tumour growth effects by antagonising the effects of HGF and its receptor, cMET, in human lung cancer cells. J Transl Med, 13: 280.
- Wang WS and Zhao CS (2021) Formononetin exhibits anticancer activity in gastric carcinoma cell and regulating miR-542-5p. Kaohsiung J Med Sci, 37: 215-25.
- Wang AL, Li Y, Zhao Q and Fan LQ (2018) Formononetin inhibits colon carcinoma cell growth and invasion by microRNA‑149‑mediated EphB3 downregulation and inhibition of PI3K/AKT and STAT3 signaling pathways. Mol Med Rep, 17: 7721-9.
- Yi L, Cui J, Wang W, et al. (2020) Formononetin Attenuates Airway Inflammation and Oxidative Stress in Murine Allergic Asthma. Front Pharmacol, 11: 533841.
- Wang H, Zhang D, Ge M, Li Z, Jiang J and Li Y (2015) Formononetin inhibits enterovirus 71 replication by regulating COX- 2/PGE₂ expression. Virol J, 12: 35.
- Cheki M, Yahyapour R, Farhood B, et al. (2018) COX-2 in Radiotherapy: A Potential Target for Radioprotection and Radiosensitization. Curr Mol Pharmacol, 11: 173-83.
- Han ZQ, Liao H, Shi F, et al. (2017) Inhibition of cyclooxygenase-2 sensitizes lung cancer cells to radiation-induced apoptosis. Oncol Lett, 14: 5959-65.
- Pan C, Zhang Y, Meng Q, Dai G, Jiang Z and Bao H (2019) Down Regulation of the Expression of ELMO3 by COX2 Inhibitor Suppresses Tumor Growth and Metastasis in Non-Small-Cell Lung Cancer. Front Oncol, 9: 363.
- Xu YQ, Long X, Han M, et al. (2021) Clinical benefit of COX-2 inhibitors in the adjuvant chemotherapy of advanced non-small cell lung cancer: A systematic review and meta-analysis. World J Clin Cases, 9: 581-601.
- Fernandes JV, Cobucci RN, Jatobá CA, Fernandes TA, de Azevedo JW and de Araújo JM (2015) The role of the mediators of inflammation in cancer development. Pathol Oncol Res, 21: 527-34.
- Wang CC, Yuan JR, Wang CF, et al. (2017) Anti-inflammatory Effects of Phyllanthus emblica L on Benzopyrene-Induced Precancerous Lung Lesion by Regulating the IL-1β/miR-101/Lin28B Signaling Pathway. Integr Cancer Ther, 16: 505-15.
- Hinshaw DC and Shevde LA (2019) The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res, 79: 4557-66.
- Russo E, Guizzardi M, Canali L, et al. (2023) Preoperative systemic inflammatory markers as prognostic factors in differentiated thyroid cancer: a systematic review and meta-analysis. Rev Endocr Metab Disord, 24: 1205-16.
- Kwon Y, Kim Y, Eom S, et al. (2015) MicroRNA-26a/-26b-COX-2-MIP-2 Loop Regulates Allergic Inflammation and Allergic Inflammation-promoted Enhanced Tumorigenic and Metastatic Potential of Cancer Cells. J Biol Chem, 290: 14245-66.
- Frejborg E, Salo T and Salem A (2020) Role of Cyclooxygenase-2 in Head and Neck Tumorigenesis. Int J Mol Sci, 21: 9246.
- Ma X, Wang R, Yu S, Lu G, Yu Y and Jiang C (2020) Anti-Inflammatory Activity of Oligomeric Proanthocyanidins Via Inhibition of NF-κB and MAPK in LPS-Stimulated MAC-T Cells. J Microbiol Biotechnol, 30: 1458-66.
- Jin J, Guo T, Guo Y, et al. (2019) Methylation‑associated silencing of miR‑128 promotes the development of esophageal cancer by targeting COX‑2 in areas with a high incidence of esophageal cancer. International journal of oncology, 54: 644-54.
- Li T, Lu J, Zhong Y (2013) Lentivirus-mediated shRNA interference targeting cyclooxygenase-2 inhibits growth of human non-small cell lung cancer. Journal of B.U.ON. : official journal of the Balkan Union of Oncology, 18: 908-14.
- Canavese M, Santo L and Raje N (2013) Cyclin dependent kinases in cancer: potential for therapeutic intervention. Cancer Biol Ther, 13: 451-7.
- Xu Y, Lv F, Zhu X, Wu Y and Shen X (2016) Loss of asparagine synthetase suppresses the growth of human lung cancer cells by arresting cell cycle at G0/G1 phase. Cancer Gene Ther, 23: 287-94.
- Cinar AS, Bakar-Ates F and Onder A (2020) Seseli petraeum M. Bieb. (Apiaceae) Significantly Inhibited Cellular Growth of A549 Lung Cancer Cells through G0/G1 Cell Cycle Arrest. An Acad Bras Cienc, 92: e20191533.
- Hua N, Wei X, Liu X, et al. (2012) A novel muscarinic antagonist R2HBJJ inhibits non-small cell lung cancer cell growth and arrests the cell cycle in G0/G1. PLoS One 7: e53170.
- Montalto FI and De Amicis F (2020) Cyclin D1 in Cancer: A Molecular Connection for Cell Cycle Control, Adhesion and Invasion in Tumor and Stroma. Cells, 9: 2648.
- Tchakarska G and Sola B (2020) The double dealing of cyclin D1. Cell Cycle 19: 163-178, 2020.
- Kobliakov VA (2019) The Mechanisms of Regulation of Aerobic Glycolysis (Warburg Effect) by Oncoproteins in Carcinogenesis. Biochemistry (Mosc), 84: 1117-28.
- Soto-Heredero G, Gómez de Las Heras MM, Gabandé-Rodríguez E, Oller J and Mittelbrunn M (2020) Glycolysis - a key player in the inflammatory response. FEBS J, 287: 3350-69.
FIGURE 1
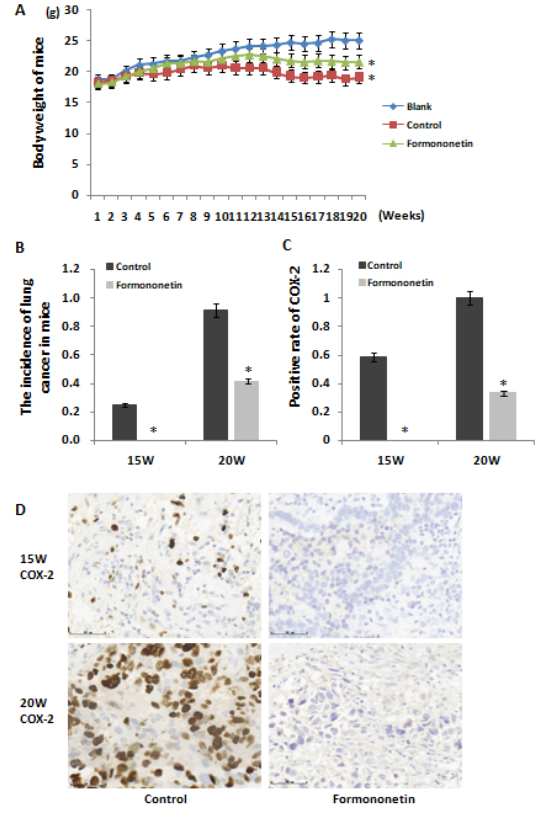
Figure 1: The formononetin treatment reduced the incidence of lung cancer and decreased the expression of COX-2 in mice. (A) The body weight of mice in each group, the mice in blank group received no treatment, in control group received urethane treatment, and in formonone group received urethane treatment and formonone treatment. (B) The incidence of lung cancer in mice at week 15 and 20 after urethane treatment. (C-D) COX-2-positive rate in lung tissues of mice at week 15 and 20 after urethane treatment.
FIGURE 2
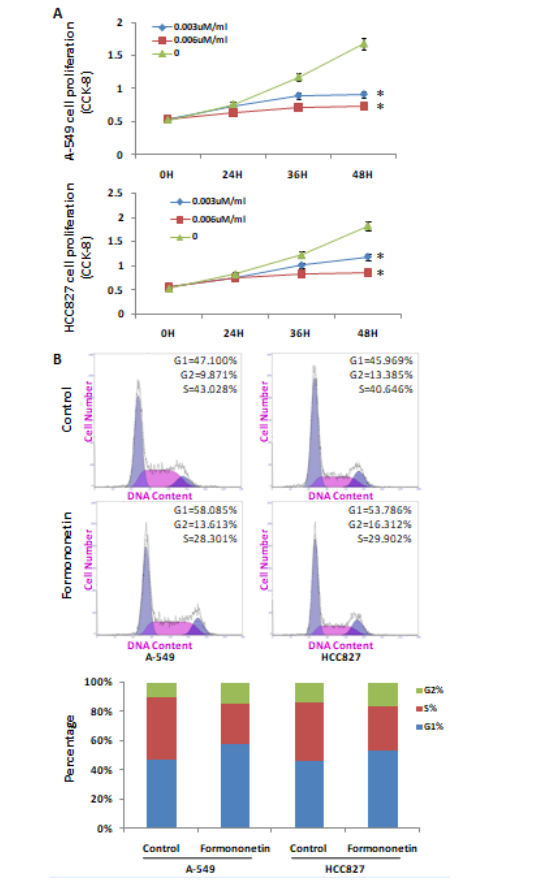
Figure 2: The role of formononetin in NSCLC cells. (A) The effect of formononetin on the viability of NSCLC cells by CCK-8. (B) The effect of formononetin on the cell cycle of NSCLC cells by flow cytometry.
FIGURE 3
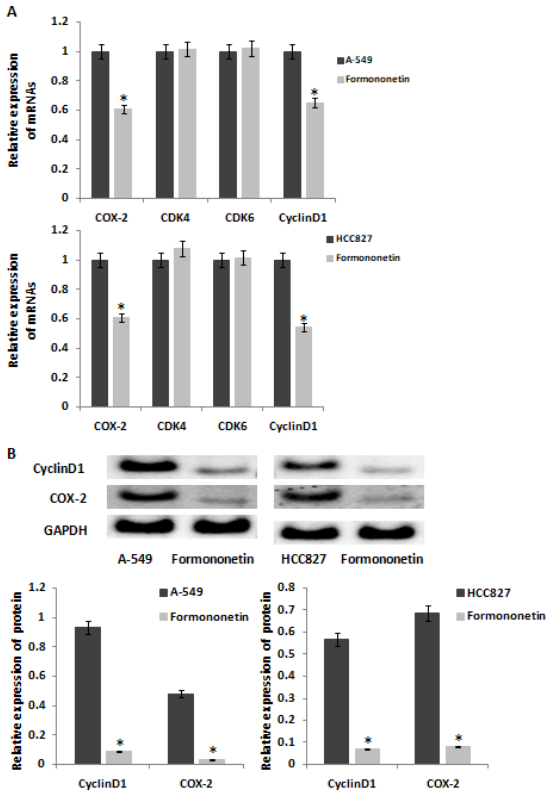
Figure 3: Formononetin inhibited the expression of COX-2 and CyclinD1. (A) The mRNA expression of CDK4, CDK6, CyclinD1 and COX-2 was tested by qRT-PCR in A-549 and HCC827 cells after formononetin treatment. (B) The protein expression of CyclinD1 and COX-2 was tested by Western blotting in A-549 and HCC827 cells after formononetin treatment.
FIGURE 4
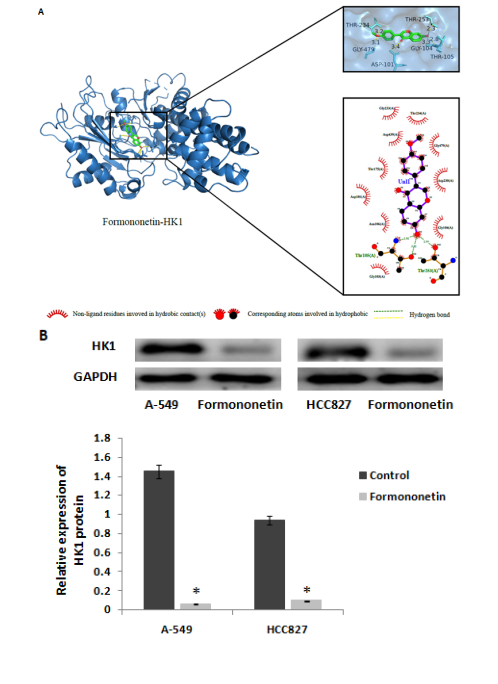
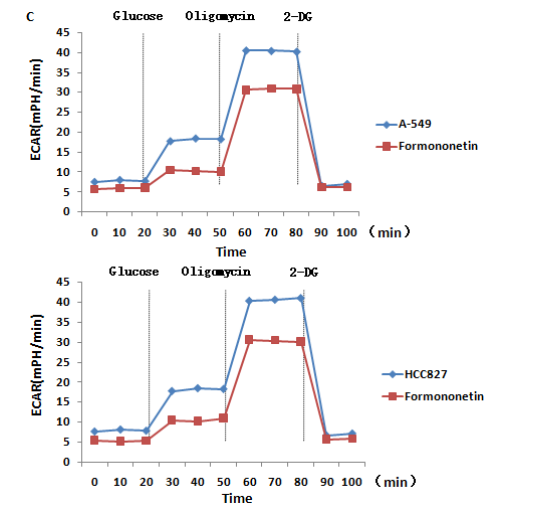
Figure 4: Formononetin targets and decreases the expression of HK1. (A) The binding site of formononetin and HK1. (B) The levels of HK1 protein were tested by Western blotting. (C) The effect of formononetin on the glycolysis rate was tested by ECAR in A-549 and HCC827 cells.
Figures at a glance