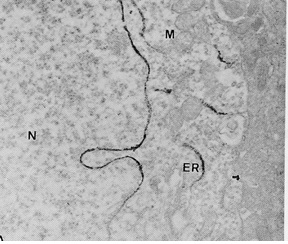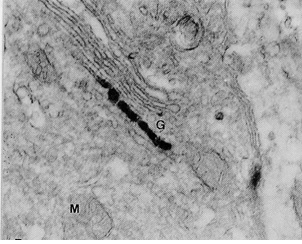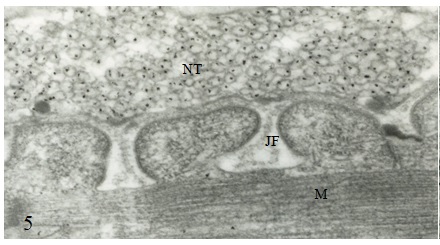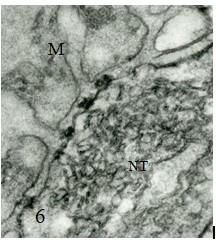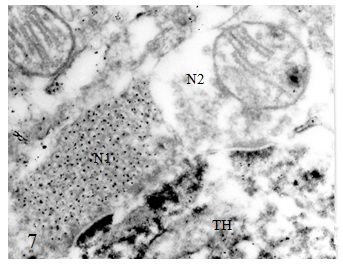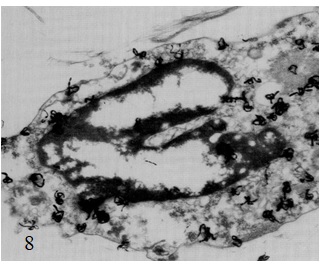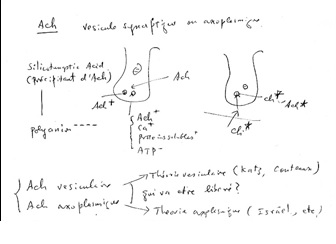
Figure 1 Draft written in French by Tsuji on the two main theories of acetylcholine (Ach) release in the cholinergic synapse. Acetylcholine is released in the synaptic cleft from exocytosis of synaptic vesicles (vesicular theory, favoured by B. Katz and R. Couteaux) or through axoplasmic diffusion (non-vesicular or axoplasmic theory, favoured by M. Israël). Still today, the mechanism of acetylcholine release remains to be clearly elucidated.