Human Papilloma Virus Genotype and DNA Methylation of TP53, PIK3CA as Integrated Biomarkers for Oro Pharyngeal Carcinogenesis, A Review
Received Date: October 18, 2023 Accepted Date: November 18, 2023 Published Date: November 21, 2023
doi: 10.17303/jocr.2023.4.203
Citation: Garba Yahaya, Efosa Bolaji Odiegie, Blessing Emosho Yemhe, Nosa Terry Omorodion (2023) Human papilloma Virus Genotype and DNA Methylation of TP53, PIK3CA as Integrated Biomarkers for Oro Pharyngeal Carcinogenesis, A Review. JJ Oncol Clin Res 4: 1-11
Abstract
Head and neck squamous cell carcinoma (HNSCC), especially oropharyngeal squamous cell carcinoma (OPSCC), has recently been found to be significantly associated with human papillomavirus (HPV) infection [1]. The incidence of OPSCC has been increasing and surpassed the number of cervical cancer cases in the United States [2]. Head and neck squamous cell carcinoma (HNSCC) encompasses a heterogeneous group of malignant neoplasms arising from the non-keratinizing epithelium of the upper aero digestive tract [3]. The aim of this review is to understand genetic roles of HPV, methylation of TP53, and PIK3CA in the development of OPSCC. Research publication articles from PubMed and many other were searched using goggle search engine. Over fifty journals of international reputes were obtained, out of which more than forty-five gives reputable results information on HPV and methylation of TP53, PIK3CA integration in OPSCC. Therefore, HPV, methylation of TP53 and PIK3CA are found to be indispensable Biomarkers in the development of OPSCC.
Keywords: HNSCC- Head and Neck Squamous Cell Carcinoma, OPSCC- Oropharyngeal squamous cell carcinoma, HPVHuman papillomavirus
Introduction
The molecular underpinnings of human papillomavirus (HPV)-related carcinogenesis in the context of head and neck squamous cell carcinoma (HNSCC) and focuses on HPV-positive oropharyngeal squamous cell carcinoma in areas for which specific data is available [4]. It covers the major pathways dysregulated in HPV-positive HNSCC and the genome-wide changes associated with this disease [5]. Head and neck squamous cell carcinoma (HNSCC) encompasses a heterogeneous group of malignant neoplasms arising from the non-keratinizing epithelium of the upper aero digestive tract [6]. Anatomic subsites of HNSCC include the oral cavity, nasopharynx, oropharynx, hypopharynx, and larynx [7]. Squamous cell carcinoma arising from these subsites collectively represents the sixth most common malignancy worldwide, accounting for 932,000 new cases and 379,000 deaths in 2015 [8].
Over the past four decades, striking epidemiological trends have been observed in HNSCC [9]. Although the overall incidence of HNSCC has declined slightly, the relative contribution of each anatomic subside to the overall incidence of HNSCC has shifted dramatically [10]. Incidence rates of tumours arising from non-oropharyngeal subsides (oral cavity, hypopharynx and larynx) have decreased while the incidence of oropharyngeal squamous cell carcinoma has steadily grown [11]. These subset-specific epidemiological trends have been attributed to shifts in societal factors that have resulted in changes due to exposure to two divergent, but complementary classes of HNSCC. risk factors :(1) tobacco and alcohol consumption and (2) human papillomavirus (HPV) infection. Successful public health campaigns in high-income countries are largely credited with achieving population-level decreases in tobacco and alcohol consumption, with concomitant declines in tobacco-associated tumours such as non-oropharyngeal HNSCC and lung cancer. Trends toward sexual practices that increase the risk of contracting sexually transmitted pathogens, like HPV, have been linked to the rise in HPV-associated cancers including oropharyngeal HNSCC (OPSCC) and anal cancers [12]. Currently, HPV-positive OPSCC cases are surpassing the incidence of HPV-positive cervical cancer [2].
Human papillomavirus is the most common sexually transmitted infection in the United States and the primary infectious cause of HNSCC [13]. Although the spread of high-risk HPV infection is pervasive, the virus is cleared by most people within 18 months [80]. It is believed that persistent infection is necessary for the development of HPV-associated malignancies [14]. The oropharynx exhibits the strongest relationship to HPV. Indeed, HPV-positive oropharyngeal squamous cell carcinoma (OPSCC), is recognized as a distinct neoplastic entity with a unique molecular, histopathological, epidemiological, and clinical profile [5]. Patients with HPV-positive OPSCC diverge from the classical profile of HNSCC patients in that they are more often younger than 60 years in age and less likely to report a history of heavy tobacco and alcohol consumption [15]. HPV-positive OPSCC also exhibits marked sensitivity to treatment and a significantly better prognosis than HPVnegative HNSCC [16]. Observations which have led to the establishment of a staging system specific to this entity [17]. and shifts in treatment paradigms. These distinct epidemiological and clinical features are manifestations of the unique underlying biology of HPV-positive OPSCC [18]. However, much of the research into the role of HPV in HNSCC was conducted before intimate links between HPV and OPSCC were widely recognized [19].
Humanpapilloma VirusPapillomaviridae is an ancient clade of non-enveloped viruses with a circular double-stranded DNA genome [20]. Within this family, approximately 200 human papillomavirus genotypes, or ‘types’, have been described based on viral genome sequence [21]. Twelve HPV types have been defined by the WHO as high-risk and exhibit high oncogenic potential [22]. At least 10 of these oncogenic HPV types (HPV16, 18, 31, 33, 45, 51, 52, 56, 58, and 59), as well as 6 low-risk HPV types (11, 32, 44, 53, 57, and 81), have been isolated from HNSCC tumours. HPV16 represents the primary viral cause of HNSCC and is identified in at least 87% of HPV-positive OPSCC [23]. HPV18 and HPV33, the next most prevalent types, account for most of the remainder of HPV-positive HNSCC [24].
All papillomaviruses possess a genome of approximately 8 kilo base encoding 8 open reading frames (ORFs) that enable viral genome replication and viral particle assembly [25]. ORFs of the HPV genome are organized based on the timing of expression with respect to the viral life cycle: early (E1, E2, E4, E5, E6, and E7) and late (L1 and L2) genes [26]. The E1 viral helicase and E2 DNA-binding protein directly mediate viral genome replication, while E4, E5, E6, and E7 are accessory proteins that coordinate viral genome amplification and virulence [27]. The late genes L1 and L2 encode viral capsid proteins necessary for the final stages of virion assembly and mediate viral entry into future host cells [28]. The functional diversity of this limited set of ORFs is expanded through complex patterns of viral transcript splicing and stage-specific gene expression that is linked to host cell differentiation [29].
Infection of HpvThe host cells for HPV infection are keratinocyte progenitors located in the basal layer of stratified squamous epithelia and adhered to the epithelial basement membrane [30]. Experimental models suggest that infection requires viral access to the basement membrane. In the epidermis and anogenital tract, HPV gains access to basal cells through micro-abrasions that occur during sexual or other direct physical contact [31].
Difference Between Humanpalloma Virus Positive Opscc and Hpv Negetive OpsccHPV status has widely been described as an independent predictor of improved overall and disease-free survival (OS and DFS) in OPSCC patients [32]. One of the first studies to demonstrate these improved outcomes of 74% reduction in risk of death from cancer among patients with HPV-related OPSCC was found compared with those with non HPV-related OPSCC [5]. A meta-analysis confirmed the data showing that patients with HPV-related OPSCC have a 28% reduce risk of death (meta HR: 0.72, 95% CI: 0.5–1) compared with non HPV-related OPSCC [33]. Several phase II and III clinical trials in locally advanced HNSCC have also shown a survival benefit for patients with HPV-related cancers [34]. In addition, the RTOG 0129 study indicates that clinical factors, such as smoking history and stage, could also influence the prognosis of patients with HPV-related HNSCC [4]. The combination of HPV status, tobacco smoking (pack-year) and tumour stage has shown to classify patients as having a low, intermediate, or high risk of death [18] 3-year rates of OS were 93% (95% CI 88.3–97.7), 70.8% (95% CI 60.7–80.8) and 46.2% (95% CI 34.7–57.5), respectively. Additional analyses have indicated that patients with p16INK4a-positive OPSCC with T4 or N2c-N3 disease have a high risk of disease progression, even if tobacco exposure is < 10 pack years [35]. It is estimated that>20% of patient with OPSCC have bad prognosis and this population need to be clearly identified [36].
Hpv Detection TechniqueHPV testing is now mandatory for an accurate diagnosis and prognosis of patients with OPSCC [37]. There are several available techniques such as p16INK4a IHC, detection of HPV-DNA by in situ hybridisation (ISH) or polymerase chain reaction (PCR), E6/E7 HPV-mRNA evaluation by ISH and reverse transcriptase– PCR (RT–PCR). Each technique yields different sensitivity and specificity profiles, but E6/E7 HPV-mRNA evaluation is considered the gold standard to confirm HPV causality since it detects oncogene transcriptional active HPVs. The p16INK4a expression is a surrogate marker of HPV involvement and it is the most widely implemented technique in the clinical setting [38]. In OPSCC, p16INK4 shows high sensitivity >90% and moderate >80% specificity compared with HPV16 E6- mRNA expression [24]. However, using p16INK4a IHC alone is questionable, because a subset of HPV-DNA and mRNAnegative HNSCCs show diffuse p16INK4a staining, indicating expression is not specific for HPV activity [76] (e.g. a mutation on Rb can also overexpress p16INK4a).
HPV Integration and HNSCCThe integration of HR-HPV DNA into the host genome has been considered an important biological step in the development of carcinogenesis in invasive cervical cancer and HNSCC [39]. Initial studies demonstrated that transcriptionally active integrated and/or episomal viral DNA in HNSCC cell lines was independent of viral copy number and integration sites. HPV integration can lead to host genomic instability, such as deletions, inversions, and chromosomal translocations [5]. A number of viral integration sites in the host genome were found in intergenic regions as well as cancer-associated genes such as TP63, ETS2, RUNX1, FOXA1 and ERBB2 [40]. Moreover, viral integration into cellular genes was commonly identified in recurrent HPV16-positive OPSCC patients, and these cellular genes are related to cancer-associated signaling pathways or mechanisms [41]. Integrated viral DNA copies could be in tandem. Viral DNA integration through the disruption of the viral E2 region leads to increased transcription of viral E6 and E7 [42]. Tumors with HPV DNA integration differ from HPV integration-negative tumors by different patterns of DNA methylation and gene expression profiles. Recently, found that HPV 16 and HPV 18 integration events in cervical cancer and HNSCC were associated with local variations and genomic rearrangements based on the Pan-- Cancer Analysis of Whole Genomes Consortium [44]. HPV integration inducing genome instability is hypothesized to be a secondary genetic event in the carcinogenesis of HPVassociated HNSCC [19]. HPV infection is associated with increased expression of the APOBEC genes APOBEC3A and APOBEC3B but exclusively with known driver genes such as TP53, CDKN2A and TERT [45]. These findings suggest a possible role of APOBECs in HPV-induced carcinogenesis, i.e., the activity of APOBECs as C-to-U RNA editing enzymes contributes to alterations in host genome expression, and APOBEC3A increases tumorigenesis in vivo [46]. In addition, as part of the immune defense system, APOBEC3A can sensitize cancer cells to cisplatin treatment by activating base excision repair and mediating the repair of cisplatin interstrand crosslinks [47]. These results suggest a role of impaired antiviral defence in driving the carcinogenesis of HPV-related HNSCC. HPV16 insertions also lead to the amplification of the PIM1 serine/threonine kinase gene in HNSCC cell lines [48]. The inhibition of PIM family kinases successfully decreased cell proliferation in vitro and in vivo in an HNSCC model [49]. Notably, viral integration can be found in both tumors that respond to treatment and recurrent tumors with more complex integration patterns in host gene [50]. By analysing viral-host fusion transcripts, showed that the HPV-positive but HPV integration negative subgroup had better survival than the HPV integration-- positive subgroup and HPV-unrelated HNSCC [51]. Moreover, HPV-positive but HPV integration-negative tumors had enhanced tumor infiltrates of immune cells and upregulated immune-related genes [5]. Consistently, another study indicated that HPV-related HNSCC can be subdivided into an immune cell enrichment phenotype and a phenotype with higher proliferation Thus, the enhanced immune profile in patients with HPV-positive but HPV integration-negative tumors may be attributed to better survival for these patients [43]. However, potential mechanisms for HPV integration-induced oncogenes [5].
Genetic and Epigenetic AlterationsPIK3CA is the most frequently mutated gene in human papillomavirus (HPV)-associated oropharyngeal squamous cell carcinoma (OPSCC). Prognostic implications of such mutations remain unknown [5]. We sought to elucidate the clinical significance of PIK3CA mutations in HPVassociated OPSCC in patients treated with definitive chemo radiation (CRT) [52].
Recent landmark sequencing studies have demonstrated gene expression profiles and somatic mutations such as TP53, CDKN2A, PTEN, PIK3CA, EGFR, HRAS, FBXW7 and NOTCH1 in diverse anatomical sites of HNSCC [53]. The Cancer Genome Atlas Network 2015). Importantly, diversity in the number of mutations and gene profiles was seen in patients with a history of tobacco use and between HPV-related and HPV-unrelated tumours [54]. The mutation rate of HPV-related tumors was almost half that of HPV-unrelated tumours [55]. Thus, two etiologies may result in the alteration of oncogenes and tumour suppressor genes that have tumorigenic effects involved in multistep biological processes [56]. HPV-related HNSCC harbours mutations in the oncogene PIK3CA encoding PI3K catalytic p110 subunit alpha, a loss of TRAF3 and the amplification of E2F1 (The Cancer Genome Atlas Network 2015) [57]. A recent comprehensive analysis on oral squamous cell carcinoma (OSCC) identified secondary genetic alterations, including PIK3CA, ZNF750 and EP300 as candidate cancer driver genes [58]. APOBEC cytosine deaminase editing was associated with genomic mutation burden in HPV-related OSCC [59]. APOBEC-mediated cytosine deamination leading to PIK3CA mutations is involved in the tumorigenesis of HPV-driven tumours [60]. As we discussed above, virushost interactions, as seen by the interaction between HPV integration with APOBEC and others, may shape genomic alterations and facilitate tumorigenesis [61]. Notably, PIK3- CA mutations (2.6% to 19%) lead to the activation of the PI3K-AKT-mTOR1 signalling pathway necessary for the viral life cycle [51]. Moreover, both HPV-related and HPVunrelated HNSCCs harbour PIK3CA mutations, and higher expression of PIK3CA in primary tumors is associated with tumor recurrence and chemo- and radio resistance [62]. Thus, inhibitors targeting the PI3K-AKT-mTOR pathway have been developed for cancer therapies [63]. However, the clinical response rates remain modest in these studies and warrant further investigation [64]. Studies in patient-derived xenograft (PDX) models demonstrate that EGFR, KT1and CSMD1 copy number aberrations are related to the effect of PI3-kinase inhibition regardless of the status of PIK3CA mutation.
Hpv and Dna MethylationHPV is a small, circular, double-stranded virus that targets the basal layer of the epithelial cells in the head and neck region, HPV targets the oropharynx, especially the tonsils, and the base of the tongue [4]. There are more than 200 HPV types, which can be divided into high-risk and low-risk types based on their potential to induce cancer [65]. Persistent high-risk HPV infections can progress to invasive cancer within 10 years, although the majority of these infections are cleared within 1 or 2 years [66]. In HNSCC, >90% of HPV-associated cases involve HPV16, which is classified as a high-risk HPV [5].
HPV16 is approximately 7900 bp in size. It exists in the nucleus of infected cells as a circular episome [67]. The proteins produced early during the infection are known as early proteins: E1, E2, E4, E5, E6, and E7 [68]. The proteins produced late during the infection are known as late proteins: L1 and L2. There is a long control region that codes no protein between the L1 stop codon and E6 AUG, and it contains the early viral promoter p97 [26]. Another promoter p670 that is related to the late viral promoter exists in the E7 coding region [26]. Based on these two promoters, HPV16 oncoproteins are generated. Among these HPV proteins, E2 inhibits the p97 promoter and results in inhibition of E6 and E7 [69]. Therefore, inhibition of E2, such as E2 disruption caused by HPV genome integration to human genome or DNA methylation of E2 binding site, causes the upregulation of E6 and E7 [5]. As mentioned herein, although HPV E6 and E7 are oncoproteins and inactivate p53 and RB respectively, these proteins also regulate the DNA methylation of the host genome [70].
For example, wild-type p53 negatively regulates DNA methyltransferase 1 (DNMT1) expression by forming a complex with specificity protein 1 (Sp.1) and chromatin modifiers on the DNMT1 promoter in lung cancer [71]. In HPV-associated HNSCC, degradation of p53 is generally caused by the HPV E6 oncoprotein and DNMT1 is consequently upregulated [19]. Additionally, the HPV E7 oncoprotein has been reported to form a complex with DNMT1 and DNMT1 is upregulated in HPV-associated OPSCC. and cervical cancer [19]. Apart from E6 and E7, c-Myc (MYC) is also reported to recruit DNA methyltransferase 3 alpha (DNMT3A). and the MYC-associated genetic network is reported to be activated in HPV-associated HNSCC [72]. The molecular mechanism of induction of DNA methylation in human papillomavirus (HPV)--associated head and neck squamous cell carcinoma (HNSCC) [72]. Wild-type p53 negatively regulates DNA methyltransferase 1 (DNMT1). HPV E6 oncoprotein cause degradation of p53 and it results in the upregulation of DNMT1 (left). HPV E7 oncoprotein forms a complex with DNMT1 and results in DNMT1 upregulation (middle) [73]. MYC is upregulated in HPV-associated HNSCC and recruit DNMT3B (right) [19].
Epigenetic Dysregulation in OPSCCThese biological differences between HPV-associated HNSCC and HPV-negative HNSCC are partly explained by the differences in mutation patterns [5]. Frequent mutations in several genes, such as TP53, CDKN2A, and PIK3CA, as well as in members of the NOTCH pathway, have been reported in HPV-negative HNSCC via genomic and transcriptomic approaches [74]. In HPV-associated OPSCC, p53 and RB are mainly inactivated by HPV E6 and E7 oncoproteins, respectively, therefore, somatic mutations in TP53 and CDKN2A are very rare [70]. However, somatic mutations in PIK3CA, E2F1, and TRAF3 have been reported, and chromatin regulators, such as lysine methyltransferase2C (KMT2C), KMT2D, CREB-binding protein (CREBBP), and E1A-associated protein p300 (EP300), are also mutated in HPV-associated OPSCC [72]. In addition, apolipoprotein B mRNA-editing catalytic polypeptide-like (APOBEC)-mediated mutagenesis, such as PIK3CA mutation and HPV genome mutation, has been reported in HPV-associated HNSCC [5]. The APOBEC3 signature is also displayed in HPV-associated HNSCC. Overall, HPV-associated OPSCC shows relatively fewer genetic alterations in cancer drivers than HPV-negative tumours at the exome level [5].
Apart from mutations, epigenetic dysregulation is also a common pathological feature in human malignancy [75]. HNSCC samples have been characterized according to their patterns of DNA methylation, one of the critical epigenetic mechanisms that silence tumour suppressor genes in cancers [76]. In particular, it has been reported that viral infections can induce aberrant DNA methylation during carcinogenesis and HPV-associated HNSCC tends to harbour a higher amount of aberrantly methylated DNA than HPVnegative HNSCC [72].
DNA Methylation Targeted TherapyEpigenetic-targeted therapy, especially targeting DNMTs, has the potential for tackling HPV-associated HNSCC [72]. 5-azacytidine and 5-aza-2′-deoxycytidine are the most used, U.S. Food and Drug Administration-approved drugs (2004 and 2006, respectively) [77]. These drugs are cytidine analogs that are incorporated into DNA, leading to covalent adduct formation and working as DNMTs inhibitors [78]. They are used only for the treatment of some myelodysplastic syndrome and chronic myelomonocytic leukemia cases, and their efficacy for solid cancers is under consideration, with clinical trials still ongoing [79]. One clinical trial is currently using 5-azacytidine for the treatment of HPV-associated and HPV-negative HNSCC. HPV-associated HNSCC tends to have higher methylation levels compare with HPV-negative HNSCC; thus, these drugs might be good candidates for treating HPV-associated HNSCC [72].
Conclusion
HPV induces DNA methylation in a complex manner during carcinogenesis. In this review, we provided an overview of DNA methylation, the relationship between DNA methylation and HPV-associated HNSCC, and how these mechanisms are related to the carcinogenesis of HPVassociated HNSCC, methylation of PIK3CA in HPV-associated OPSCC and Non HPV-associated OPSCC, and methylation of TP53 in HPV-associated OPSCC and Non HPV related OPSCC. Recent comprehensive large-cohort DNA methylation analyses at a genome-wide scale have revealed that there is an HPV-associated HNSCC subtype with increased DNA methylation. However, there is still room for elucidation of the mechanism of HPV and DNA methylation. A more detailed understanding of the molecular basis of this subtype might lead to the development of new therapeutic strategies, such as therapeutic de-escalation in this subtype.
- Lifsics A, Cistjakovs M, Sokolovska L, Deksnis R, Murovska M et al. (2023) The Role of the p16 and p53 Tumor Suppressor Proteins and Viral HPV16 E6 and E7 Oncoproteins in the Assessment of Survival in Patients with Head and Neck Cancers Associated with Human Papillomavirus Infections. Cancers, 15: 2722.
- Ndon S, Singh A, HA PK, Aswani J, Chan JY-K et al. (2023) Human Papillomavirus-Associated Oropharyngeal Cancer: Global Epidemiology and Public Policy Implications. Cancers, 15: 4080.
- Jagadeeshan S, Novoplansky OZ, Cohen O, Kurth I, Hess J et al. (2023) New insights into RAS in head and neck cancer. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer, 188963.
- Ferris RL and Westra W (2023) Oropharyngeal carcinoma with a special focus on hpv-related squamous cell carcinoma. Annual Review of Pathology: Mechanisms of Disease, 18: 515-35.
- Lim YX, Mierzwa ML, Sartor MA and D’silva NJ (2023) Clinical, morphologic and molecular heterogeneity of HPV-associated oropharyngeal cancer. Oncogene, 1-17.
- Sivasakthivel S, Ramani P, Krishnan RP and Krishnan SR, RP (2023) Systematic Review and Meta-Analysis on Angiotensin Converting Enzyme 2 in Head and Neck Region. Cureus, 15.
- Abuei H, Namdari S, Pakdel T, Pakdel F, Andishe-- Tadbir, (2023) Human parvovirus B19 infection in malignant and benign tissue specimens of different head and neck anatomical subsites. Infectious Agents and Cancer, 18: 1-10.
- Tofanelli M (2020) Application of Nanotechnology To The Diagnosis And Treatment Of Oral And Oropharyngeal Cancer.
- Yang T-H, Xirasagar S, Cheng Y.-F, Chen C.-S, Chang W.-P et al. (2023) Trends in the incidence of head and neck cancer: A nationwide population-based study. Oral Oncology, 140: 106391.
- Bedard MC, Chihanga T, Carlile A, Jackson R, Brusadelli MG et al. (2023) Single cell transcriptomic analysis of HPV16-infected epithelium identifies a keratinocyte subpopulation implicated in cancer. Nature Communications, 14: 1975.
- Mazul AL, Chidambaram S, Zevallos JP and Massa ST (2023) Disparities in head and neck cancer incidence and trends by race/ethnicity and sex. Head & Neck, 45: 75-84.
- Davison K (2023) Risk Factors of Human Papillomavirus Positive Oropharyngeal Cancer in Men. Walden University.
- Chaudhary V, Chaudhary N, Mathews S and Singh RD (2023) Human Papilloma Virus− Associated Oral Pharyngeal Squamous Cell Carcinoma: Prevalence, Prevention, and Awareness of Vaccination in the Indian Population. Critical Reviews™ in Oncogenesis, 28.
- Kusakabe M, Taguchi A, Sone K, Mori M and Osuga Y (2023) Carcinogenesis and management of human papillomavirus-associated cervical cancer. International Journal of Clinical Oncology, 1-10.
- Brennan S (2023) Human Papillomavirus Associated Oropharyngeal Squamous Cell Carcinoma: Genetic Aberrations Associated with Local and Distant Recurrence.
- Wang R, Khurram SA, Walsh H, Young LS and Rajpoot N (2023) A novel deep learning algorithm for human papillomavirus infection prediction in head and neck cancers using routine histology images. Modern Pathology, 100320.
- Huber S, Baer C, Hutter S, Dicker F, Meggendorfer M et al. (2023) AML classification in the year 2023: How to avoid a Babylonian confusion of languages. Leukemia, 1-8.
- Ahn D, Kwak, J.-H, Lee G.-J and Sohn J.-H (2023) Prevalence and Characteristics of Human Papillomavirus Infection in Oropharyngeal Squamous Cell Papilloma. Cancers, 15: 810.
- Chakraborty S, Ramasubbu K, Banerjee M, Balaji MP, Vinayagam, Y et al. (2023) A systematic review on the molecular and clinical association between Human Papillomavirus and Human Immunodeficiency Virus co‐infection in Head, Neck and Oral squamous cell carcinoma. Reviews in Medical Virology, e2462
- Coronado PLG, Sáenz CIR, Kawas JR, Triviño DGZ, Zayas YR et al. (2023) Design, synthesis, and evaluation of peptides derived from L1 protein against bovine papillomavirus-1/2 identified along Mexico’s cattle export route. Journal of Veterinary Research.
- Yao X, Xu Z, Duan C, Zhang Y, Wu X et al. (2023) Role of human papillomavirus and associated viruses in bladder cancer: An updated review. Journal of Medical Virology, 95: e29088.
- Gong X, ChI H, Xia Z, Yang G and Tian G (2023) Advances in HPV‐associated tumor management: Therapeutic strategies and emerging insights. Journal of Medical Virology, 95: e28950.
- Zhu Y, He H, Zhu X, Hu Y, Yu S et al. (2023) Epidemiology of oral human papillomavirus infection among diverse Chinese adults in typical areas of China: Findings from the DLCC study. Cancer Epidemiology, Biomarkers & Prevention.
- Andrioaie IM, Luchian I, Dămian C, Nichitean G, Andrese E P (2023) The Clinical Utility of Circulating HPV DNA Biomarker in Oropharyngeal, Cervical, Anal, and Skin HPV-Related Cancers: A Review. Pathogens, 12: 908.
- Andrioaie IM, Luchian I, Dămian C, Nichitean G, Andrese E P (2023) The Clinical Utility of Circulating HPV DNA Biomarker in Oropharyngeal, Cervical, Anal, and Skin HPV-Related Cancers: A Review. Pathogens, 12: 908.
- Jönsson J (2023) The balance of splicing: A novel insight into the splicing regulation of high-risk HPV E6 and E7 oncogenes. Acta Universitatis Upsaliensis
- Ludwig CH, Thurm AR, Morgens DW, Yang KJ, Tycko J et al. (2023) High-throughput discovery and characterization of viral transcriptional effectors in human cells. Cell Systems, 14, 482-500. e8.
- Ryabchenko B, Šroller V, Horníková L, LovtsoV A, Forstová J et al. (2023). The interactions between PML nuclear bodies and small and medium size DNA viruses. Virology Journal, 20: 1-20.
- Gimenez J, SpallonI A, Cappelli S, Ciaiola F, Orlando V et al. (2023) TDP-43 Epigenetic Facets and Their Neurodegenerative Implications. International Journal of Molecular Sciences, 24, 13807.
- Yin W, EgawA N, Zheng K, GriffiN H, Tian P et al. (2023) HPV E6 inhibits E6AP to regulate epithelial homeostasis by modulating keratinocyte differentiation commitment and YAP1 activation. PLoS Pathogens, 19: e1011464.
- Trüeb RM and Dutra Rezende H (2023) Viral Diseases of the Hair and Scalp. Hair in Infectious Disease: Recognition, Treatment, and Prevention. Springer.
- Oka T, Sato F, Ono T, Kawaguchi T, Murotani K et al. (2023) Prognostic values of systemic inflammation and nutrition‐based prognostic indices in oropharyngeal carcinoma. Laryngoscope Investigative Otolaryngology
- Volpe S, Gaeta A, Colombo F, Zaffaroni M, Mastroleo F et al. (2023) Blood-and Imaging-Derived Biomarkers for Oncological Outcome Modelling in Oropharyngeal Cancer: Exploring the Low-Hanging Fruit. Cancers, 15: 2022.
- Roof L and Yilmaz E (2023) Immunotherapy in HPV-Related Oropharyngeal Cancers. Current Treatment Options in Oncology, 24: 170-83.
- Santaló Corcoy M (2020) De-escalation of the adjuvant radiotherapy doses in patients with HPV-related oropharyngeal tumors with high levels of CD8+ tumor infiltrating lymphocytes: a multicentre, randomized, controlled clinical trial.
- Farber NI, LI, Y, Solis RN, Chen J, Masheeb Z Ea al. (2023) Tumor and Nodal Disease Growth Rates in Patients with Oropharyngeal Squamous Cell Carcinoma. Cancers, 15: 3865.
- Paolini F, Campo F, Iocca O, Manciocco V, De Virgilio A et al. (2023) It is time to improve the diagnostic workup of oropharyngeal cancer with circulating tumor HPV DNA: Systematic review and meta‐analysis. Head & Neck.
- Li R, Xiao L, Gong T, Liu J, Li Y et al. (2023a). Role of oral microbiome in oral oncogenesis, tumor progression, and metastasis. Molecular Oral Microbiology, 38: 9-22.
- Chen B, Zhao L, Yang R And Xu T (2023) Advances in molecular mechanism of HPV16 E5 oncoprotein carcinogenesis. Archives of Biochemistry and Biophysics, 109716.
- Patrizi S (2022) Multi-omics approaches to complex diseases in children.
- Li X, Ling Y, Hu L, Zhang L, Lin S (2023b) Detection of HPV DNA, E6/E7 mRNA, and p16INK4a in lung cancer: A systematic review and meta-analysis. The Journal of Infectious Diseases, jiad295.
- Thiruvengadam, R and Kim JH (2023) Therapeutic strategy for oncovirus-mediated oral cancer: A comprehensive review. Biomedicine & Pharmacotherapy, 165: 115035.
- Qin T, Li S, Henry Le, Chou E, Cavalcante RG et al. (2023) Whole-genome CpG-resolution DNA Methylation Profiling of HNSCC Reveals Distinct Mechanisms of Carcinogenesis for Fine-scale HPV+ Cancer Subtypes. Cancer Research Communications, 3, 1701-1715.
- Rodriguez I, Rossi NM, Keskus A, Xie Y, Ahmad T et al. (2023) Insights into the Mechanisms and Structure of Breakage-Fusion-Bridge Cycles in Cervical Cancer using Long-Read Sequencing. medRxiv 21: 23294276.
- Xian S, Dosset M, Castro A, Carter H and Zanetti M (2023) Transcriptional analysis links B cells and TERT expression to favorable prognosis in head and neck cancer. PNAS nexus, 2: pgad046.
- Warren Cj, Santiago Ml, Pyeon D (2022) APOBEC3: friend or foe in human papillomavirus infection and oncogenesis? Annual Review of Virology, 9: 375-95.
- Petrilla C, Galloway J, Kudalkar R, Ismael A, Cottini F (2023) Understanding DNA Damage Response and DNA Repair in Multiple Myeloma. Cancers, 15: 4155.
- Wei Y, Zhao Z, Ma X (2022) Description of CRISPR-Cas9 development and its prospects in human papillomavirus-driven cancer treatment. Frontiers in Immunology 13: 1037124.
- Liu W, Qaed E, Zhu Y, Tian W, Wang Y et al. (2023) Research Progress and New Perspectives of Anticancer Effects of Emodin. The American Journal of Chinese Medicine, 1-43.
- Jassim A, Rahrmann EP, Simons BD, Gilbertson RJ (2023) Cancers make their own luck: theories of cancer origins. Nature Reviews Cancer, 1-15.
- Dong H, Shu X, Xu Q, Zhu C, Kaufmann AM et al. (2021) Current status of human papillomavirus-related head and neck cancer: from viral genome to patient care. Virologica Sinica 36: 1284-302.
- Bhatia A, Burtness B (2023) Treating head and neck cancer in the age of immunotherapy: A 2023 update. Drugs 83: 217-48.
- Park Hy, Lee Js, Wee Jh, Kang Jw, Kim Es et al. (2023) Assessment of the Mutation Profile of Tonsillar Squamous Cell Carcinomas Using Targeted Next-Generation Sequencing. Biomedicines 11: 851.
- 54. Eldridge RC, Qin ZS, Saba NF, Houser MC, Hayes DN et al. (2023) Unsupervised Hierarchical Clustering of Head and Neck Cancer Patients by Pre-Treatment Plasma Metabolomics Creates Prognostic Metabolic Subtypes. Cancers 15: 3184.
- Saba NF, Pamulapati S, Patel B, Mody M, Strojan P et al. (2023) Novel Immunotherapeutic Approaches to Treating HPV-Related Head and Neck Cancer. Cancers, 15: 1959.
- Latha, HA, Kommalapati V, Naag S, Kalyan S, Vidya, KS et al. (2023) Role of Virus in Oncogenesis–A Systemic Review.
- Van Der Kamp MF, Halmos GB, Guryev V, Horvatovich PL, Schuuring E et al. (2022) Age-specific oncogenic pathways in head and neck squamous cell carcinoma-are elderly a different subcategory? Cellular Oncology 45: 1-18.
- Patten VA (2023) The analysis of genetic aberrations in South African oesophageal squamous cell carcinoma patients.
- Kleszcz R (2023) Advantages of the Combinatorial Molecular Targeted Therapy of Head and Neck Cancer—A Step before Anakoinosis-Based Personalized Treatment. Cancers 15: 4247.
- Schrank TP, Kothari A, Weir WH, Stepp WH, Rehmani H et al. (2023) Noncanonical HPV carcinogenesis drives radiosensitization of head and neck tumors. Proceedings of the National Academy of Sciences 120: e2216532120.
- Qualliotine JR, Nakagawa T, Rosenthal SB, Sadat S, Ballesteros-Merino C et al. (2023) A Network Landscape of HPVOPC Reveals Methylation Alterations as Significant Drivers of Gene Expression via an Immune-Mediated GPCR Signal. Cancers, 15: 4379.
- Kostopoulou ON, Zupancic M, Pont M, Papin E, Lukoseviciute M et al. 2022. Targeted Therapy of HPV Positive and Negative Tonsillar Squamous Cell Carcinoma Cell Lines Reveals Synergy between CDK4/6, PI3K and Sometimes FGFR Inhibitors, but Rarely between PARP and WEE1 Inhibitors. Viruses 14: 1372.
- Jin J, He J, Li X, Xiaoqi, N, Jin X (2023) The role of ubiquitination and deubiquitination in PI3K/AKT/mTOR pathway: a potential target for cancer therapy. Gene, 147807.
- Goodman RS, Jung S, Balko JM, Johnson DB (2023) Biomarkers of immune checkpoint inhibitor response and toxicity: challenges and opportunities. Immunological Reviews.
- Hinton H, Coleman S, Salem J, Kingsley K (2023) Screening for High-Risk Oral Human Papillomavirus (HPV31, HPV33, HPV35) in a Multi-Racial Pediatric and Adult Clinic Patient Population. Cancers 15: 4501.
- Reuschenbach M, Doorbar J, Del Pino M, Joura EA, Walker C (2023) Prophylactic HPV vaccines in patients with HPV-associated diseases and cancer. Vaccine.
- Rossi NM, Dai J, Xie Y, Wangsa D, Heselmeyer-Haddad K et al. (2023) Extrachromosomal Amplification of Human Papillomavirus Episomes Is a Mechanism of Cervical Carcinogenesis. Cancer Research 83: 1768-81.
- Chen SY, Last A, Ettyreddy A, Kallogjeri D, Wahle B et al. (2021) 20 pack-year smoking history as strongest smoking metric predictive of HPV-positive oropharyngeal cancer outcomes. American journal of otolaryngology 42: 102915.
- Evande R, Rana A, Biswas-Fiss EE, Biswas SB (2023) Protein–DNA Interactions Regulate Human Papillomavirus DNA Replication, Transcription, and Oncogenesis. International Journal of Molecular Sciences 24: 8493.
- Kassab A, Gupta I, A Moustafa Ae (2023) Role of E2F transcription factor in oral cancer: Recent insight and advancements. Seminars in Cancer Biology
- Cirino G, Szabo C, Papapetropoulos A (2023) Physiological roles of hydrogen sulfide in mammalian cells, tissues, and organs. Physiological Reviews 103: 31-276.
- Nakagawa T, Kurokawa T, Mima M, Imamoto S, Mizokami H et al. 2021. DNA methylation and HPV-associated head and neck cancer. Microorganisms 9: 801.
- Trejo Cerro O (2023) Regulation of the Human Papillomavirus E7 Oncoprotein. The Open University
- Huang X, Duijf PH, Sriram S, Perera G, Vasani S et al. (2023) Circulating tumour DNA alterations: emerging biomarker in head and neck squamous cell carcinoma. Journal of biomedical science 30: 1-19.
- Bhattacharya S, Mahato RK, Singh S, Bhatti GK, Mastana SS et al. (2023) Advances and challenges in thyroid cancer: The interplay of genetic modulators, targeted therapies, and AI-driven approaches. Life Sciences 122110.
- Kylmä AK (2023) HPV-related and unrelated oropharyngeal cancer: Contribution of oral pathobionts Treponema denticola and Porphyromonas gingivalis to carcinogenesis. Dissertationes Scholae Doctoralis Ad Sanitatem Investigandam Universitatis Helsinkiensis.
- Scholpa NE (2023) Role of DNA methylation during recovery from spinal cord injury with and without β2-adrenergic receptor agonism. Experimental Neurology 368: 114494.
- Laranjeira AB, Hollingshead MG, Nguyen D, Kinders RJ, Doroshow JH, Yang SX (2023) DNA damage, demethylation and anticancer activity of DNA methyltransferase (DNMT) inhibitors. Scientific Reports 13: 5964.
- Molica M, Perrone, S, Rossi M (2023) Immunotherapy of Acute Myeloid Leukemia: A Work in Progress.
- Doorbar J (2023) The human Papillomavirus twilight zone–Latency, immune control and subclinical infec- tion. Tumour Virus Research 200268.
FIGURE 1
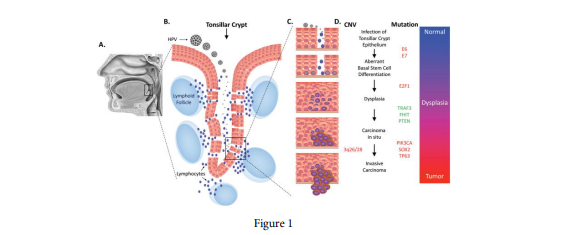
Figure 1:
Figures at a glance