Chondrocytes: From their Origin to their Application in Regeneration of Articular Cartilage
Received Date: October 28, 2023 Accepted Date: November 28, 2023 Published Date: November 30, 2023
doi: 10.17303/jscr.2023.5.102
Citation: Karla Mendez-Maldonado, Itzel Escobedo-Avila, Sara Caballero-Chacon (2023) Chondrocytes: From their origin to their application in regeneration of articular cartilage. J Stem Cell Rep 5: 1-17
Abstract
Understanding the mechanisms that regulate chondrogenesis and cartilage development will contribute to detection of diseases and disorders that affect cartilage and will provide foundation for novel prevention and treatment approaches, such as gene therapy and tissue engineering. Cellular therapy based on chondrocytes implantation is the most widely used procedure for inducing cartilage regeneration. In this review, we will discuss about the different source of chondrocytes (mesenchymal stem cells, adipose stem cells, blood cells and neural crest), the chondrocytes-derived disease, the scaffolds that are using in medicine field to treat the articular cartilage injury and the animal models to study this lesion as well as the cartilage regeneration. We propose use the neural crest cells to prove these cells in the different scaffolds to articular regeneration.
Keywords: Chondrocytes; Neural Crest; Regeneration; Scaffolds; Articular Cartilage; Stem Cell
Introduction
Embryonic origin and chondrocyte differentiation
Cartilage is a connective tissue that attends multiple prenatal and postnatal functions and is formed mostly by cells called chondrocytes. Chondrocytes exhibit a life cycle of proliferation, differentiation, maturation, and apoptosis and the rate of each of these processes is dependent on temporal and spatial cues within the body [1]. The origins of the cartilage lineage are diverse; chondrocytes arise from cranial neural crest cells (CNCCs) of the neural ectoderm, cephalic mesoderm, sclerotome of the paraxial mesoderm, or somatopleure of the lateral plate mesoderm. Terminal differentiation of chondrocytes results in different types of cartilage: hyaline, elastic and fibrous. The formation of cartilage is initially characterized by aggregation of chondrogenic progenitor mesenchymal cells into nodules. This condensation process is dependent on cell-matrix, cell-cell adhesion signals and cytoskeletal architecture. During this process, cells group at sites of future skeletal elements in response of multiple growth and differentiation factors and morphogens such as Wnts, the transforming growth factor superfamily (TGFβs), including TGFβs, bone morphogenetic proteins (BMPs), and fibroblast growth factors (FGFs) in the extracellular environment [2]. Although chondrogenesis is regulated by combinatorial signaling of a great number of factors, cell condensation is the major event of cell commitment to the cartilage lineage, after which tissue-specific transcription factors and structural proteins begin to accumulate.
At this stage, MSCs express type I and type II A collagens, which are nonspecific for chondrocytes [3]. Type II A procollagen, one of the two splice forms of type II procollagen, is present in human pre-chondrogenic and newly formed cartilage tissues [4,5]. High expression of fibronectin, hyaluronic, CD44, neural cadherin and neural cell adhesion molecules has also been detected at these sites, as well as numerous intracellular signaling pathways transduced by integrin’s, mitogen-activated protein kinases, protein kinase C, lipid metabolites and cAMP [2]. These elements suggest that aggregation and condensation of chondrocytes are mediated by both cell-cell and cell-matrix adhesion [6-8].
In growth cartilage, approximately 50% of the total volume of tissue is cellular. The chondrocytes are characterized by a high potential for cell division, forming several columns of proliferative cells. The activity of proliferation is associated with a renewal of replacement cells by the recruitment of pre-chondrocytes. Chondrocytes reach their highest level of metabolic activity in this phase and secrete matrix molecules until each cell is included in a dense matrix divided into two zones: a peripheral area which encompasses and extends over the articular surface, and a central area which also serves as the center of the endochondral ossification of the epiphysis. The tissue becomes recognizable as cartilage when matrix accumulation separates the cells, and they take a spherical shape.
Members of the FGF family (2,4,8,9,10 and 18), together with the FGF receptors (1,2 and 3) coordinate patterning and cell proliferation during chondrogenesis and endochondral ossification in embryonic and postnatal growth plates. Sox9 is also expressed by proliferative chondrocytes in growth cartilage and induces the expression of components of cartilage matrix, including collagen type II in vitro [9].
Chondrogenic differentiation occurs under the influence of factors still poorly defined. It involves the maturation of pre-chondrocytes into mature chondrocytes, which is characterized by the extinction of the genes encoding type I collagen and cellular adhesion molecules, and the expression of a few matrix proteins specific to chondrocytes, including type IX and XI collagens, and proteoglycans (PGs) such as aggrecan. The type II collagen mRNA splice form switches from type IIA to IIB procollagen, the major ECM component of cartilage [4].
Some factors involved in chondrogenic differentiation have been characterized in mice and mammals, such as the growth factors FGF-2, 4 and 8 [10,11], members of the TGFβ and BMP families [12,13] and transcription factors of the Sox family, Sox9, L-Sox5 and Sox6 [14,15].
Chondrocytes are the only cell type constituting the cartilage, a connective tissue found in synovial joints, articular discs, nose, and ears of mammals. They synthesize the cartilage matrix (collagen, proteoglycans, and elastin fibers) and maintain homeostasis in articular regions. Chondrocytes comprise 2% of the total volume of articular cartilage [16].
There are various types of cartilage, depending on the age of the individual. Thus, chondrocytes have different phenotypes depending on the growth stage or age of the individual. Additionally, they differ in size, shape, and metabolic activity in different areas of cartilage. The functional specificity of the different types of cartilage depends on the degree of cell proliferation and maturation, the nature of proteins synthesized and the three-dimensional arrangement of these proteins around the cell [17].
Sox9 promotes chondrocyte differentiation by activating chondrocyte-specific enhancer elements in type II, IX, XI collagens and aggrecan genes [18]. Moreover, Sox9 deletion in undifferentiated limb bud MSCs results in a complete failure of chondrogenesis, and if this gene is inactivated following initiation of chondrogenesis, most collagen type I-expressing cells fail to undergo to the differentiation stage [19].
In the final stage of development, the chondrocytes at central zone secrete proteins that assist in calcification of the matrix, whereas chondrocytes present at peripheral zones synthesize and release collagen and other ECM molecules to develop hyaline cartilage. The mature chondrocytes remain recruited within ECM, are rounded, and possess limited regeneration potential [20]. The chondrocytes only represent 1%–2% of total volume of articular cartilage and play key role in synthesizing the degraded ECM to maintain its functional nature [21]. The cellular density of chondrocytes is highest in early ages (0–2 years) and decreases with the advancing age of individual [22]. The shape, size, and number of chondrocytes vary according to zones and regions of articular cartilage [21]. In superficial zones, they are flat and small. The density of chondrocytes in deeper zones is less as compared to superficial zone. Chondrocytes secrete ECM and are immobilized within this specific environment, which affects the turnover of ECM. This entrapment inhibits direct cell-to-cell interaction. However, the cells respond to stimuli such as growth factors, mechanical loads, and hydrostatic pressure [23]. It has also been reported that chondrocytes contain a cell-autonomous circadian rhythm, the disruption of which was observed in osteoarthritic individuals [24].
How to obtain chondrocytes?
The limited ability of regeneration of chondral tissue upon loss has led to look for solutions to reconstruct damaged or lost tissues. The use of stem cells has become a focus on stem cell-based therapy for cartilage defects due to its pluripotency, abundant sources and low immunogenicity. Currently, there are various sources of stem cells for cartilage generation in vitro.
Mesenchymal Stem Cells
Mesenchymal stem cells (MSCs) are multipotent cells with a broad proliferative potential and ability to differentiate into various cell types, such as osteocytes, adipocytes, chondrocytes, myocytes, cardiomyocytes and neurons [25]. These cells are characterized by the expression of numerous surface antigens, but none of them appears to be exclusively expressed on MSC. The main source of mesenchymal stem cells is the bone marrow; however, they only constitute a small percentage of bone marrow populating cells. MSCs play a key role in the maintenance of bone marrow homeostasis and regulate the maturation of both hematopoietic and non-hematopoietic cells. MSC have become the most extensively used stem cells in regenerative medicine because they can be derived from a variety of sources (bone marrow, adipose tissue, muscle, periodontal ligament, and synovial fluid), have low immunogenicity, no ethical concerns, and a minimal teratoma risk formation. Moreover, it has been reported that these cells engraft successfully in patients and cause beneficial effects [26].
Adipose Stem Cells
Adipose-derived stem cells (ASCs) are a subset of MSCs that can be easily obtained from adipose tissues and possess many of the same regenerative properties as other MSCs. ASCs have become one of the most promising stem cell populations identified so far because adipose tissue is relatively abundant in the human body and therefore, ASCs can be obtained in higher numbers with less donor-site morbidity. ASCs harvested from different anatomical areas exhibit different characteristics; isolated from different locations, cell types, and species have different density and different procedures may affect quality, functionality, and plasticity [27]. They are a good source of cells for treatment of cartilage lesions due to ease of their collection and their capacity for multipotential differentiation. One gram of adipose tissue yields approximately 5000 stem cells, whereas the yield from bone marrow derived mesenchymal stem cells is 100 to 1000 cells/mL of marrow [28]. Although ASCs are of mesodermal origin, it has been shown that they can differentiate into cells of ectodermal, endodermal, and mesodermal origin. These are the second most used stem cells in cartilage tissue engineering, which can be isolated in high numbers with minimal manipulation and can be expanded in vitro without loss of their chondrogenic potential. Several works have used adipose-derived stem cells to differentiate into functional and mature chondrocytes, using different procedures, such as mechanical stress or small molecules which has been reported to induce the up regulation of Sox9 and promote chondrogenic phenotypes [29,30]. Recently, it has been reported the use of electrical stimulation to differentiate adipose-derived stem cells to chondrocytes without the addition of any exogenous factors [31].
Peripheral Blood Mononuclear Cells (PBMSCs)
Peripheral blood is a large accessible source of adult stem cells for both basic research and clinical applications. Peripheral blood mononuclear cells (PBMCs) have been reported to contain distinct multipotent progenitor cell populations and possess the potential to differentiate into blood cells, endothelial cells, hepatocytes, cardiomyogenic cells, smooth muscle cells, osteoblasts, osteoclasts, epithelial cells, neural cells, or myofibroblasts under appropriate conditions. Furthermore, transplantation of these PBMC-derived cells can regenerate tissues and restore function after injury. Blood is the most convenient source from which to obtain SCs from patients with minimal invasiveness, and can be frozen and stored for later use [32]. Recently PBMCs can be easily extracted from peripheral blood with minimal invasiveness. Recently, PBMCs have been reported to be able to undergo chondrogenic differentiation and have a cartilage generation ability like that of MSCs [33].
Human Embryonic Stem Cells (hESC)
Embryonic stem cells (ESC) are pluripotent cells which give rise to all somatic cell types in an organism. ESC can be a valuable tool for understanding mechanisms involved in development of specialized cells and establishment of organ structures. Moreover, the indefinite self-renewal ability and plasticity of ESC allows the in vitro generation of distinct cell types for regenerative medicine. The greatest therapeutic promise of hESC is to generate specialized cells to replace damaged tissue in patients suffering from various degenerative diseases [34]. hESC have the potential to differentiate it to chondrocytes. These cells have been used to expand chondrocytes under selective growth factors and cultured in hyaluronic acid (HA)-based hydrogel.These cells had been used to regenerate osteochondral tissue in an osteochondral defect model using rats [35].
Neural Crest Cells
Another source of chondrocytes recently defined is the neural crest stem cells. Neural crest (NC) cells are a migratory cell population synonymous with vertebrate evolution. Neural crests are located between the neural plate and the neural ectoderm, forming a fourth germ layer. They generate a wide variety of cell and tissue types during embryonic and adult development including cartilage and bone, connective tissue, pigment and endocrine cells as well as neurons and glia among other phenotypes [36-41]. Recently, the neural crest has been proposed like a new source of chondrocytes to cartilage regeneration, since neural crest cells can be easy to expand in culture dish and these cells can be isolate from different accessible source like nasal cartilage, ear perichondrium tissue, oral tissues [42-47].
Our research group set the goal of determining in our previous mESC transgenic lines and H1 hESC could be differentiated to chondrocytes neural crest derived using different protocols reported previously [48,49]. We evaluated by RT-qPCR pluripotency marker expression in mouse cell lines and human cell lines as well as induction neural crest markers and chondrocytes differentiation markers after differentiation protocol. We used mESC from previous work to get some preliminary results to neural crest induction and differentiation [40].
How we mentioned above, there are many sources to chondrocytes differentiation, (mesenchymal cells, blood cells, adipose cells, and neural crest cells) could differentiate to chondrocytes but the efficiency to generate enough chondrocytes to use in implantation treatment or articular regeneration is poor.For this reason, some scientist groups are working on increase the chondrocytes proliferation using scaffolds (as we discuss below), small molecules [50], chemokines and glycoside like Salidroside. Salidroside seems has an interesting characteristic to increase the chondrocytes proliferation.In this work, the authors demonstrated that this compound, who was obtained from a root of Rhodiola Rosea and has been reported inhibited the pro-inflammatory responses in osteoarthritis, increased chondrocytes proliferation in a rat model [51].
Chondrocytes and Diseases
Osteoarthritis
Osteoarthritis (OA) is the most common form of arthritis, characterized by progressive cartilage degradation but also synovial membrane inflammation [52,53].
Mesenchymal progenitor cells initially condensate and rapidly differentiate into chondrocytes.Phenotypic modification of chondrocytes is an important mechanism that contributes to the loss of cartilage in OA [54,55]. The pathogenesis of OA remains unclear, and it cannot effectively prevent the progression of OA. During OA onset, chondrocytes modify their synthesis activity [56].
The acquisition of hypertrophic phenotype by chondrocytes is among the most documented one and constitute a central contributing factor for OA pathogenesis and other degenerated phenotypes at the onset of OA such as fibro-chondrocytes including the dedifferentiated-like phenotype [57-59].
Kashin-Beck Disease
Is an endemic, chronic and degenerative osteoarthropathy principally occurring in children? Its main clinical symptoms include diarthrodial joint deformation and limited joint mobility this disease is characterized by chondrocytes necrosis in hyaline articular cartilage, matrix destruction, resulting in the loss of proteoglycans (PGs) [60]. PGs are one of the major components in the extracellular matrix (ECM) of articular cartilage. Aggrecan is one of the major PGs in the ECM of articular cartilage. The function of articular cartilage is dependent on a high aggrecan concentration being present in the tissue [19]. In degenerative joint disease such as OA and RA, loss of PGs, like Aggrecan, is the signal in disease onset and development [61].
Rheumatoid Arthritis
Rheumatoid arthritis (RA) is a chronic autoimmune syndrome, affecting the articular joints contributing to cartilage and bone damage. Etiology of this disease is not clear, several immune pathways, involving immune cells (T cells, B cells, dendritic cells, macrophages and neutrophils) and nonimmune (fibroblast and chondrocytes) cells, participate in the secretion of pro-inflammatory proteins and matrix lysing enzymes that could change the immune balance and promote to cartilage and bone damage [62]. In RA, multiple cytokines produced by inflammatory cells, decrease viability and proliferation of chondrocytes [63]. On the other hand, evidence also implicated chondrocytes as effector cells in RA directly and indirectly, by releasing multiple enzymes of ECM degradation, as metallo proteases, pro inflammatory proteins promote aggrecanase production [64].
Animal models to study Chondrogenesis Disease
We can obtain chondrocytes from different sources, but it is necessary to test it in some animal models to help us to understand if these chondrocytes can proliferate, migrate and if they can integrate in and be part of the tissue after of implantation. For this reason, there are several works, where the authors used different strategies to identify the best source and the best conditions to have the best and efficient implant. In case of articular cartilage it is important to mention that this tissue was believed to not have immunogenic properties, for this reason they usually treat the Chondrogenic disease using allogenic transplantation.
Autologous chondrocyte implantation (ACI) and scaffold supported ACI are used for treatment of larger chondral defects (>2 cm2). These implicate an open surgery re implantation of ex-vivo cultured autologous chondrocytes harvested as a biopsy. This procedure can fail to regenerate articular cartilage and need reoperation [65], because the articular cartilage has immunogenic properties and needs be modulated [66,67]. For this reason, it is necessary to investigate new methods for optimizing the chondrocytes, growth and use the best scaffold to support the chondrocytes.
In the next section we review the current sources to obtain more chondrocytes and the methodologies to increase the cartilage repair using scaffolds, small molecules and growth factors.
Rabbit
Rabbit is an animal model that is widely used in research to study different types of diseases, rehabilitation, and stem cell therapy [68-71], in case of implantation research not widely used to evaluate directly the efficacy of human chondrocytes implantation.
One of the transplantations used in rabbits to study articular regeneration is using the autologous or allogenic chondrocytes. In 2019, Boopalan et al. demonstrated that regeneration after chondrocytes transplantation autologous or allogenic form was similar, the transplantation generated hyaline/mixed hyaline cartilage and they did not observe significant differences [72].
In 2018, Takahashi et al., administered an immunosuppressant tacrolimus, at different doses, to use human chondrocytes sheets to transplant into knee osteochondral defects in Japanese white rabbits.They evaluate at different times the possible rejection of the transplant. They decided that in the higher doses of immunosuppressor it is possible to use the rabbit as a preclinical model for the evaluation of human cell-based therapies [73]. However, the research to use this model animal continues.
In 2019, Zheng et al., investigated if osteochondral regeneration is possible using another type of scaffold, they used three-dimensional printed polycaprolactone-hydroxyapatite (PCL-HA) scaffold coated with umbilical cord blood mesenchymal stem cells (UCB-MSCs) and chondrocytes. The authors showed that the UCB chondrocytes-seeded PCL-HA scaffolds promote articular cartilage repair demonstrating that UCB seeded-chondrocytes promote bone and cartilage regeneration [74]. Using the scaffolds technology and the cell therapy is possible to improve the articular regeneration, in this case a study in 2020 showed that using allogenic mesenchymal stem cells and using growth factors like IGF-1 (insulin like growth factor-1) or TGF-b (transforming growth factor b-1) accelerated osteochondral healing [75].
Mouse
There are many reports about the cartilage regeneration using different strategies, like the regeneration by activation of stem cells close to the chondral injury. In this work, the authors activated the skeletal stem cells (SSC) using microfracture in knee joints, they showed that the resident-stem cells population in the injury site can be induced to generate cartilage and this cartilage were used for treatment of the chondral injury. They found that the combination of BMP2 (a potent osteogenic growth factor) and VEGF inhibition using soluble sVEGFR1 promote the cartilage formation in resting SSC. This combination promotes expression of cartilage markers in vitro and in vivo [76]. Other report using hydrogels-based and anti-miRNAs showed that it is possible to increase cartilage regeneration using endogenous cells, they used anti-miR-221 and this were transfected into human bone marrow-derived mesenchymal stromal cells (hMSC), this blockade the expression of miR-221 and resulted an increase in chondrogenesis, they used the 3D pellets technology and developed hydrogel-based system for delivering anti-miR-221 to endogenous cells in situ using the hMSC, then they evaluated chondrogenic markers like Col2 and GAGs by immunostainings and showed an increase in these markers in osteochondral biopsies that were implanted subcutaneously in mice [77].
On the other hand, the use of small molecules to promote the cartilage regeneration has taken great importance since Kartogenin was using to enhanced chondrogenic differentiation [78] and induce cartilage regeneration in osteoarthritis but KGN can be cleaved into 4-aminobipheny (4-ABP) and phthalic acid (PA) and these peptides were tested in a mouse model of osteoarthritis, the administered the peptides via oral and intra-articular and evaluated if they can promote the cartilage regeneration too. The authors found that the 4-ABP was more efficient than the KGN because 4-ABP was found in cartilage and induce proliferation and chondrogenic differentiation of MSC and was able to repair the osteoarthritic lesions [79].
Recently, exosomes secreted by mesenchymal cells have been used in Osteoarthrithis therapy.But the effect of exosomes on pain relief in OA animals had not been studied. Exosomes can be endocytosed with the chondrocytes in vitro, with this idea, Lei He and co-workers used bone marrow mesenchymal stem cells (BMSC)-derived exosome to inject in cartilage damage and evaluated the pain relief and cartilage repair.They demonstrated that exosome treatment can promote cartilage repair as well as alleviate pain the OA rats [80].
Bovines
Another animal model to study the cartilage repair and scaffold integration are the bovines. In 2013, Pretzel and co-workers, analyzed the bovine cartilage regeneration in a punch model, the culture of the cartilage with and without chondrogenic factors. They used a biosynthesis of bacterial cellulose hydrogel like a scaffold for the chondrocytes. They found that the punch model is reproducible and a good tool for the long-term culture of cartilage, maintaining the integrity and homeostasis [81]. The punch model was used to evaluate the cartilage regeneration using a collagen type I hydrogel. The authors demonstrated that this hydrogel can preserve the integrity too and increased the cartilage differentiation for a long time of cultivation, up to 12 weeks [82]. Recently, has been used the resorbable polyglycolic acid (PGA) chondrocyte grafts have been used inside cartilage rings punched out of bovine femoral trochlea. The authors found that chondrocytes in PGA quickly colonized the damage site and deposition of aggrecan. After 84 days, they observed cartilage regeneration [83]. These approaches can be useful to evaluate different scaffolds to repair the damaged human cartilage. Shengu-Chuan Lin and co-workers evaluated the regenerative effects of MSC in the repair of knee cartilage defects.They use the puncture technique in the lateral femoral condyle in both knees’ joints. One week later, they treated with MSC and placebo. They observed that human adipose-derived MSC were more effective in repairing cartilage than pig umbilical cord-derived MSCs [84]. However, this must have to be demonstrated with more samples and evaluated for longer follow-up periods to verify that MSC in cartilage repair is safety and efficacy of these therapies in animals and humans.
We showed different type of models to study the chondrocytes proliferation and cartilage regeneration. In case of the rabbit model, the authors used allogenic chondrocytes or mesenchymal stem cells to increase the cartilage regeneration, but in mouse there are different type of studies where the authors used mesenchymal stem cells and different combination of biomolecules, hydrogels, or osteogenic inhibitors to increase the cartilage regeneration or by the mechanical induction like the micro fracture to induce the cartilage regeneration. On the other hand, bovine’s models used punch model and scaffolds to increase the mesenchymal stem cells derived-cartilage differentiation and deposition of aggrecan. At this point, all the animals’ models used mesenchymal stem cells or chondrocytes to increase the cartilage regeneration, using different molecules or scaffolds. The results of those works are encouraging, but the implant needs to be prolonged to apply in the different chondrocytes-derived disease. On the other hand, we need to improve to be able to use it in humans and have some mechanism for releasing molecules or stem cells specific site that allow the maintenance of the implant.
Scaffolds used for Articular Cartilage Regeneration
Chondrocytes are used for cartilage tissue repair. Chondrocytes need an environment to guide cellular function and need to be seeded in some type of biomaterial to increase the survival and promote chondrocytes to join the site of the damage. Different scaffolds have been developed to increase chondrocyte survival generating a suitable microenvironment, increase implant half-life and decrease the cost of the surgery. We will give a brief description of the type of scaffolds and in what types of surgery have been used.
Silk-based scaffolds are one of the scaffolds used to reconstruct osteochondral defects; however, there have been different types of scaffolds with different mechanical properties. In this case, the authors used Bombyx mori silkbased composite scaffold with great mechanical properties and porosity. Bombyx mori silk-based composite scaffold was fabricated embedding the natural degummed silk fibers within a matrix of regenerated fibroin. This scaffold is highly porous and have more desired mechanical specifications (ultimate tensile strength, yields pint and stiffness compared to fibroin). This scaffold has suitable degradation rate and regenerative capacity. The authors used this scaffold for 36 weeks in osteochondral lesions of rabbit knees. The efficiency of this silk-scaffold was compared with fibrin glue (FG) as a natural carrier of chondrocytes. After 36 weeks of implantation, the defects were reconstructed by chondrocytes-seeded silk scaffolds (SCF) were hyaline-like cartilage. They considered it safe and suitable for in vivo repair of osteochondral defects The repair ability of chondrocytes-seeded scaffolds is typically higher than scaffolds alone. The connective tissue was developed through embedding the natural fibers within the matrix, the authors suggest that the natural silk fibers may have a role in increasing the biodegradability of the regenerated silk matrix. The fibers increased the number of immigrated cells within the regenerated silk matrix [85]. Fibrin glue had been used in several animal studies and even human trials, but the efficiency in osteochondral disease (OCD) repair remains controversial [86-90]. FG alone or in combination with some growth factors is not useful to promote the repair of OCD [86-90], but the combination with chondrocytes or stem cells showed better approach for remodeling and repair [87-89]. One of the limitation of FG compare with silk-based composite scaffold is the short duration of evaluation time or no data about mechanical analysis to prove the efficiency of FG for OCD.
On the other hand, other types of scaffolds are Silk fibroin (SF) and Silk fibroin with gelatin/chondroitin sulfate/hyaluronate (SF-GCH). These scaffolds have been used to seed the scaffolds with human bone marrow mesenchymal stromal cells (BM-MSCs) or chondrocytes.They applied compression and evaluated the expression of aggrecan and collagen X (chondrocytes markers). They found that chondrocytes and BM-MSCs cultured in SF-GCH and compression increased glycosaminoglycan (GAG)GAG, therefore chondrocytes biosynthesis and matrix accumulation [91].
Additionally, to repair osteochondral plugs, they developed a bioprinting system using Poly (Ethylene glycol) Di methacrylate (PEGDMA) and they created three-dimensional cartilage tissue layer-by-layer. The PEGDMA were printed with human chondrocytes. They observed that printed human chondrocytes increased 26% when simultaneously polymerized and the printed in 3D bio paper had higher GAG glycosaminoglycan (GAG) content than without bio paper. They found an increase in Collagen type II and aggrecan gene expression of printed cell-hydrogel constructs in osteochondral plugs [92].
In 2019, Dufour et al, in their quest to recreate a suitable environment for chondrocytes in vitro and develop an appropriate support for hydrogel-based tissue engineering of nasal cartilage, the authors tested the capacity of IEIK13 self-assembling peptide (SAP) to serve as a support to form such cartilage gel.The human nasal chondrocytes were amplified with FGF-2 and insulin, and re-differentiated in IEIK13 with BMP-2, insulin and T3. The chondrocytes displayed round morphology and expressed cartilage-specific markers (Collagen type II and type IX and aggrecan). These SAP IEIK13 could be a suitable support for hydrogel-based tissue engineering of nasal cartilage [93].
Another type of scaffold is collagen, in this case, the authors used Type II/I collagen scaffold from rabbits and rats. They mixed Collagen with rabbit chondrocytes to fabricate neocartilage constructs in 3D systems to form neorabbit cartilage and neo-rat cartilage. The neocartilage was implanted into defects (5-mm circular and 4 mm deep) in rabbit and rat knee condyles. After 2 or 3 months, they observed through histology images cartilage-like repaired tissue, integration and healthy tissue [94].
Other trial used autologous nasal chondrocytes to repair articular cartilage in a rabbit model. They obtained neural crest in 3D alginate hydrogel and increased chondrogenic markers (Col2A1, ACAN, SOX9), the authors obtained hyaline-like repaired tissue and the mechanical analysis showed similar mechanical properties to the native cartilage such. They proposed that nasal chondrocytes can be a good seed cell source for cartilage tissue and the alginate hydrogel as delivery system [95].
Another type of implant that was recently developed was to use collagen-seeded autologous chondrocytes.In an equine model it was shown that the implantation of autologous chondrocytes improves cartilage healing and composite histologic scores, as well as increased in chondrocyte predominance, they evaluated the toluidine blue-stained matrix, and collagen type-II content compared with control. They demonstrated that this implant is quick to insert, give chondrocytes security in the defect and improves cartilage healing compared with autologous chondrocyte implantation [96].
Other types of scaffolds of natural origin have been developed such as human Warton´s Jelly extracellular matrix as biomimetic scaffold. The authors used this scaffold to promote cell repopulation and support the formation of new cartilage tissue. They found that this type of scaffold was a good candidate to treat cartilage defects in rabbits, but it is necessary to explore the application in clinical area [97].
Atelocollagen is a highly purified pepsin-treated type I collagen from calf dermis. This collagen complexed with siRNA is resistant to nucleases and is efficiently transduced into cells [98]. This type of collagen allowed longterm gene silencing site-specific and was used liked a scaffold to grow chondrocytes. This type of scaffold can be used with minced cartilage to repair large cartilage defects.The rabbit chondrocytes from minced cartilage can migrate and proliferate in atelocollagen gel in vitro. These chondrocytes embedded showed good cartilage repair equivalent to autologous chondrocyte implantation [99].
Pyrocarbon (PyC) is a synthetic material developed in the 1950s for the nuclear industry. PyC is biocompatible with blood, and it has been used in the medical field for different types of orthopedic implants [100]. Another type of biomaterial to use in implants is with CobaltChromium (CoCr).These types of material were proposed to use to mimic either cartilage-like or bone-like conditions in the context of shoulder surgical replacement. The authors use these biomaterials to evaluate the murine primary chondrocytes grown on disc made of PyC or CoCr . They demonstrated that chondrocytes grew on both types of materials, but when they analyzed the mechanical results, they observed that PyC has more elasticity and more stiffness than CoCr. They suggested that PyC is suitable for spherical interposition implants [101]. In the case of CoCr is not the best option to use in implants because it was demonstrated that this type of material is toxic, generating neurocognitive and depressive deficits after implant [102].
Conclusion
In this review, we tried to show the different chondrocytes-source types possible. We found several types of methodologies trying to increase chondrogenesis in different type of animal models as well as using different types of cells to improve the articular cartilage regeneration in the different chondrogenic diseases. We need more studies for longer-time periods to establish the best source of cells, scaffolds and/or small molecules to get a specific treatment to improve the different chondrogenic disease treatment.
The most common source of cells studied are the mesenchymal stem cells, but there is another source of chondrocytes that has been more studied recently. Neural Crest Cells (NCC) as mentioned before, is a good source of different types of cells as well as chondrocytes. This type of cells needs more studies in field chondrogenesis disease-derived therapy, but some reports showed that NCC could be a good source of chondrocytes. The combination with the different biomaterials as scaffolds and small molecules could improve the chondrocyte proliferation, differentiation and maturation, site specific using the NCC.
Acknowledgement
We would like to thanks to National Autonomous University of Mexico (Universidad Nacional Autonoma de Mexico, UNAM) postdoctoral fellowship program of Direccion General de Asuntos del Personal Academico (DGAPA) for its support during postdoctoral stay of Dr. Mendez-Maldonado.
- Shum L, Nuckolls G (2001) The life cycle of chondrocytes in the developing skeleton. Arthritis Res Ther 4.
- Steinert AF et al. (2007) Major biological obstacles for persistent cell-based regeneration of articular cartilage. Arthritis Res Ther 9: 213.
- Archer CW, Francis-West P (2003) The chondrocyte. Int J Biochem Cell Biol 35: 401-4.
- Sandell LJ, Nalin AM, Reife RA (1994) Alternative splice form of type II procollagen mRNA (IIA) is predominant in skeletal precursors and non-cartilaginous tissues during early mouse development. Developmental Dynamics 199: 129-40.
- Valcourt U, Gouttenoire J, Aubert-Foucher E, Herbage D, Mallein-Gerin F (2003) Alternative splicing of type II procollagen pre-mRNA in chondrocytes is oppositely regulated by BMP-2 and TGF-β1. FEBS Lett 545: 115-9.
- Kulyk WM, Upholt WB, Kosher RA (1989) Fibronectin gene expression during limb cartilage differentiation. Development 106: 449-55.
- Oberlender SA, Tuan RS (1994) Expression and functional involvement of N-cadherin in embryonic limb chondrogenesis. Development 120: 177-87.
- DeLise AM, Fischer L, Tuan RS (2000) Cellular interactions and signaling in cartilage development. Osteoarthritis Cartilage 8: 309-34.
- Lefebvre V, Smits P (2005) Transcriptional control of chondrocyte fate and differentiation. Birth Defects Res C Embryo Today 75: 200-12.
- Cohn MJ, Izpisúa-Belmonte JC, Abud H, Heath JK, Tickle C(1995) Fibroblast growth factors induce additional limb development from the flank of chick embryos. Cell 80: 739-46.
- Hall BK, Miyake T (2000) All for one and one for all: condensations and the initiation of skeletal development. Bio Essays 22: 138-47.
- Duprez D et al. (1996) Overexpression of BMP-2 and BMP-4 alters the size and shape of developing skeletal elements in the chick limb. Mech Dev 57: 145-57.
- Pizette S, Niswander L (2000) BMPs Are Required at Two Steps of Limb Chondrogenesis: Formation of Prechondrogenic Condensations and Their Differentiation into Chondrocytes. Dev Biol 219: 237-49.
- de Crombrugghe B et al. (2000) Transcriptional mechanisms of chondrocyte differentiation. Matrix Biology 19: 389-94.
- Bridgewater LC, Lefebvre V, de Crombrugghe B (1998) Chondrocyte-specific Enhancer Elements in the Col11a2 Gene Resemble the Col2a1 Tissue-specific Enhancer. Journal of Biological Chemistry 273: 14998-5006.
- Zylinska B et al. (2021) Structure and pathologies of articular cartilage. In Vivo 35: 1355-63.
- Akkiraju H, Nohe A (2015) Role of chondrocytes in cartilage formation, progression of osteoarthritis and cartilage regeneration. Journal of Developmental Biology 3: 177-92.
- Lefebvre V, Behringer RR, de Crombrugghe B (2001) L-Sox5, Sox6 and Sox9 control essential steps of the chondrocyte differentiation pathway. Osteoarthritis Cartilage 9: 69-75.
- Kiani C, Chen L, Wu YJ, Yee AJ, Yang BB (2002) Structure and function of aggrecan. Cell Res 12: 19-32.
- Kinner B, Capito RM, Spector M (2005) Regeneration of Articular Cartilage. in 91-123.
- Stockwell RA (1978) Chondrocytes. J Clin Pathol Suppl (R Coll Pathol) 12: 7-13.
- Mitrovic D, Quintero M, Stankovic A, Ryckewaert A (1983) Cell density of adult human femoral condylar articular cartilage. Joints with normal and fibrillated surfaces. Lab Invest 49: 309-16.
- Buckwalter JA, Rosenberg LC (1988) Electron microscopic studies of cartilage proteoglycans. Electron Microsc Rev 1: 87-112.
- Doody KM, Bottini N (2015) Chondrocyte clocks make cartilage time-sensitive material. Journal of Clinical Investigation 126: 38-9.
- Pittenger MF et al. (1999) Multilineage Potential of Adult Human Mesenchymal Stem Cells. Science 284: 143-7.
- Bobis S, Jarocha D, Majka M (2006) Mesenchymal stem cells: characteristics and clinical applications. Folia Histochem Cytobiol 44: 215-30.
- Si Z et al. (2019) Adipose-derived stem cells: Sources, potency, and implications for regenerative therapies. Biomedicine & Pharmacotherapy 114: 108765.
- Strem BM et al. (2005) Multipotential differentiation of adipose tissue-derived stem cells. Keio J Med 54: 132-41.
- Jang Y, Jung H, Ju JH (2017) Chondrogenic Differentiation Induction of Adipose-derived Stem Cells by Centrifugal Gravity. Journal of Visualized Experiments.
- Lee J et al. (2020) Differentiation of adipose-derived stem cells into functional chondrocytes by a small molecule that induces Sox9. Exp Mol Med 52: 672-81.
- Nosoudi N et al. (2021) Differentiation of adipose-derived stem cells to chondrocytes using electrospraying. Sci Rep 11: 24301.
- Zhang M, Huang B (2012) The multi-differentiation potential of peripheral blood mononuclear cells. Stem Cell Res Ther 3: 48.
- Hopper N et al. (2015) Peripheral blood derived mononuclear cells enhance osteoarthritic human chondrocyte migration. Arthritis Res Ther 17: 199.
- Vazin T, Freed WJ (2010) Human embryonic stem cells: Derivation, culture, and differentiation: A review. Restorative Neurology and Neuroscience 28: 589-603.
- Toh WS et al. (2010) Cartilage repair using hyaluronan hydrogel-encapsulated human embryonic stem cell-derived chondrogenic cells. Biomaterials 31: 6968-80.
- Simões-Costa, M. & Bronner, M. E. Insights into neural crest development and evolution from genomic analysis.Genome Res 23: 1069-80.
- Bronner ME, LeDouarin NM (2012) Development and evolution of the neural crest: An overview. Dev Biol 366: 2-9.
- Mayor R, Theveneau E (2013) The neural crest. Development 140: 2247-51.
- Mizuseki K et al. (2003) Generation of neural crestderived peripheral neurons and floor plate cells from mouse and primate embryonic stem cells. Proceedings of the National Academy of Sciences 100: 5828-33.
- Méndez-Maldonado K, Vega-López G, Caballero-Chacón S, Aybar M, Velasco I (2018) Activation of Hes1 and Msx1 in Transgenic Mouse Embryonic Stem Cells Increases Differentiation into Neural Crest Derivatives. Int J Mol Sci 19: 4025.
- Méndez-Maldonado K, Vega-López GA, Aybar MJ, Velasco I (2020) Neurogenesis From Neural Crest Cells: Molecular Mechanisms in the Formation of Cranial Nerves and Ganglia. Front Cell Dev Biol 8.
- Fulco I et al. (2014) Engineered autologous cartilage tissue for nasal reconstruction after tumour resection: an observational first-in-human trial. The Lancet 384: 337-46.
- Mumme M et al. (2016) Nasal chondrocyte-based engineered autologous cartilage tissue for repair of articular cartilage defects: ?an observational first-in-human trial. The Lancet 388: 1985-94
- Kobayashi S et al. (2011) Presence of Cartilage Stem/Progenitor Cells in Adult Mice Auricular Perichondrium. PLoS One 6: e26393.
- Togo T et al. (2006) Identification of cartilage progenitor cells in the adult ear perichondrium: utilization for cartilage reconstruction. Laboratory Investigation 86: 445-57.
- Taïhi I, Nassif A, Isaac J, Fournier BP, Ferré F (2019) Head to Knee: Cranial Neural Crest-Derived Cells as Promising Candidates for Human Cartilage Repair. Stem Cells Int 1-14.
- Méndez-Maldonado K, Vega-López G, Caballero-Chacón S, Aybar M, Velasco I (2018) Activation of Hes1 and Msx1 in Transgenic Mouse Embryonic Stem Cells Increases Differentiation into Neural Crest Derivatives. Int J Mol Sci 19: 4025.
- Kawaguchi J, Mee PJ, Smith AG (2005) Osteogenic and chondrogenic differentiation of embryonic stem cells in response to specific growth factors. Bone 36: 758-69.
- Lach MS et al. (2019) Chondrogenic differentiation of pluripotent stem cells under controllable serum-free conditions. Int J Mol Sci 20.
- Li T et al. (2020) Small molecule compounds promote the proliferation of chondrocytes and chondrogenic differentiation of stem cells in cartilage tissue engineering. Biomedicine & Pharmacotherapy 131: 110652.
- Sun M, Lu Z, Cai P, Zheng L, Zhao J (2020) Salidroside enhances proliferation and maintains phenotype of articular chondrocytes for autologous chondrocyte implantation (ACI) via TGF-β/Smad3 Signal. Biomedicine & Pharmacotherapy 122: 109388.
- Mathiessen A, Conaghan PG (2017) Synovitis in osteoarthritis: current understanding with therapeutic implications. Arthritis Res Ther 19: 18.
- Sophia Fox AJ, Bedi A, Rodeo SA (2009) The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health: A Multidisciplinary Approach 1: 461-8.
- Fukui N, Purple CR, Sandell LJ (2001) Cell biology of osteoarthritis: The chondrocyte’s response to injury. Curr Rheumatol Rep 3: 496-505.
- Sandell LJ, Aigner T (2001) Articular cartilage and changes in Arthritis: Cell biology of osteoarthritis. Arthritis Res Ther 3: 107.
- Charlier E et al. (2019) Chondrocyte dedifferentiation and osteoarthritis (OA). Biochem Pharmacol 165: 49-65.
- Aigner T, Soder S, Gebhard PM, McAlinden A, Haag J (2007) Mechanisms of Disease: role of chondrocytes in the pathogenesis of osteoarthritis-structure, chaos and senescence. Nat Clin Pract Rheumatol 3: 391-9.
- Ripmeester EGJ, Timur UT, Caron MMJ, Welting TJM (2018) Recent Insights into the Contribution of the Changing Hypertrophic Chondrocyte Phenotype in the Development and Progression of Osteoarthritis. Front Bioeng Biotechnol 6.
- Pitsillides AA, Beier F (2011) Cartilage biology in osteoarthritis—lessons from developmental biology. Nat Rev Rheumatol 7: 654-63.
- Cao J et al. (2008) Articular cartilage metabolism in patients with Kashin–Beck Disease: an endemic osteoarthropathy in China. Osteoarthritis Cartilage 16: 680-8.
- Verma P, Dalal K (2011) ADAMTS-4 and ADAMTS-5: Key enzymes in osteoarthritis. J Cell Biochem 112: 3507-14.
- Fang Q, Zhou C, Nandakumar KS (2020) Molecular and Cellular Pathways Contributing to Joint Damage in Rheumatoid Arthritis. Mediators Inflamm 3830212.
- Schuerwegh AJ et al. (2003) Influence of pro-inflammatory (IL-1α, IL-6, TNF-α, IFN-γ) and anti-inflammatory (IL-4) cytokines on chondrocyte function. Osteoarthritis Cartilage 11: 681-7.
- Tortorella MD, Malfait AM, Deccico C, Arner E (2001) The role of ADAM-TS4 (aggrecanase-1) and ADAMTS5 (aggrecanase-2) in a model of cartilage degradation. Osteoarthritis Cartilage 9: 539-52.
- Pareek A et al. Long-Term Outcomes after Autologous Chondrocyte Implantation: A Systematic Review at Mean Follow-Up of 11.4 Years. Cartilage 7: 298-308.
- Osiecka-Iwan A, Hyc A, Radomska-Lesniewska DM, Rymarczyk A, Skopinski P (2018) Antigenic and immunogenic properties of chondrocytes. Implications for chondrocyte therapeutic transplantation and pathogenesis of inflammatory and degenerative joint diseases. Central European Journal of Immunology 43: 209-19.
- Neunaber C et al. (2022) Towards the development of osteochondral allografts with reduced immunogenicity. J Mech Behav Biomed Mater 133: 105359.
- Kamaruzaman NA, Kardia E, Kamaldin N (2013) ‘Atikah, Latahir, A. Z. & Yahaya, B. H. The Rabbit as a Model for Studying Lung Disease and Stem Cell Therapy. Biomed Res Int 1-12.
- Jin ES et al. Bilateral ovario-hysterectomy induced osteoporotic rabbit model. J Biol Regul Homeost Agents 33: 391-6.
- Woodruff-Pak DS, Trojanowski JQ (1996) The older rabbit as an animal model: Implications for Alzheimer’s disease. Neurobiol Aging 17: 283-90.
- Fong IW et al. (1997) Rabbit model for Chlamydia pneumoniae infection. J Clin Microbiol 35: 48-52.
- Boopalan PR JVC, Varghese V, Sathishkumar S, Arumugam S, Amarnath V (2019) Similar regeneration of articular cartilage defects with autologous & allogenic chondrocytes in a rabbit model. Indian Journal of Medical Research 149: 650.
- Takahashi T et al. (2018) Rabbit xenogeneic transplantation model for evaluating human chondrocyte sheets used in articular cartilage repair. J Tissue Eng Regen Med.
- Zheng P, Hu X, Lou Y, Tang K (2019) A Rabbit Model of Osteochondral Regeneration Using Three-Dimensional Printed Polycaprolactone-Hydroxyapatite Scaffolds Coated with Umbilical Cord Blood Mesenchymal Stem Cells and Chondrocytes. Medical Science Monitor 25: 7361-69.
- Gugjoo MB et al. (2020) Allogeneic mesenchymal stem cells and growth factors in gel scaffold repair osteochondral defect in rabbit. Regenerative Med 15: 1261-75.
- Murphy MP et al. (2020) Articular cartilage regeneration by activated skeletal stem cells. Nat Med 26: 1583-92.
- Lolli A et al. (2019) Hydrogel-based delivery of antimiR-221 enhances cartilage regeneration by endogenous cells. Journal of Controlled Release 309: 220-30.
- Jing H et al. (2019) Kartogenin preconditioning commits mesenchymal stem cells to a precartilaginous stage with enhanced chondrogenic potential by modulating JNK and β--catenin-related pathways. FASEB J 33: 5641-53.
- Zhang S et al. (2019) Kartogenin hydrolysis product 4-aminobiphenyl distributes to cartilage and mediates cartilage regeneration. Theranostics 9: 7108-21.
- He L et al. (2020) Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res Ther 11: 276.
- Pretzel D et al. (2013) A novel in vitro bovine cartilage punch model for assessing the regeneration of focal cartilage defects with biocompatible bacterial nanocellulose. Arthritis Res Ther 15: R59.
- Horbert V et al. (2019) In Vitro Analysis of Cartilage Regeneration Using a Collagen Type I Hydrogel (CaReS) in the Bovine Cartilage Punch Model. Cartilage 10: 346-63.
- Horbert V et al. (2021) In Vitro Cartilage Regeneration with a Three-Dimensional Polyglycolic Acid (PGA) Implant in a Bovine Cartilage Punch Model. Int J Mol Sci 22: 11769
- Lin SC et al. (2023) Regenerative Effect of Mesenchymal Stem Cell on Cartilage Damage in a Porcine Model. Biomedicines 11: 1810.
- Kazemnejad S et al. (2016) Comparative repair capacity of knee osteochondral defects using regenerated silk fiber scaffolds and fibrin glue with/without autologous chondrocytes during 36 weeks in rabbit model. Cell Tissue Res 364: 559-72.
- Vogt S et al. (2009) The influence of the stable expression of BMP2 in fibrin clots on the remodelling and repair of osteochondral defects. Biomaterials 30: 2385-92.
- Berninger MT et al. (2013) Treatment of osteochondral defects in the rabbit’s knee joint by implantation of allogeneic mesenchymal stem cells in fibrin clots. J Vis Exp.
- Haleem AM et al. (2010) The clinical use of human culture-expanded autologous bone marrow mesenchymal stem cells transplanted on platelet-rich fibrin glue in the treatment of articular cartilage defects: A pilot study and preliminary results. Cartilage 1: 253-61.
- Konst YE et al. (2012) Treatment of severe osteochondral defects of the knee by combined autologous bone grafting and autologous chondrocyte implantation using fibrin gel. Knee Surgery, Sports Traumatology, Arthroscopy 20: 2263-9.
- Brittberg M, Sjogren-Jansson E, Lindahl A, Peterson L (1997) Influence of fibrin sealant (Tisseel®) on osteochondral defect repair in the rabbit knee. Biomaterials 18: 235-42.
- Sawatjui N, Limpaiboon T, Schrobback K, Klein T (2018) Biomimetic scaffolds and dynamic compression enhance the properties of chondrocyte- and MSC-based tissue-engineered cartilage. J Tissue Eng Regen Med 12: 1220-9.
- Cui X, Breitenkamp K, Finn MG, Lotz M, D’Lima DD (2012) Direct human cartilage repair using three-dimensional bioprinting technology. Tissue Eng Part A 18: 1304-12.
- Dufour A et al. (2019) Combination of bioactive factors and IEIK13 self-assembling peptide hydrogel promotes cartilage matrix production by human nasal chondrocytes. J Biomed Mater Res A 107, 893–903 (2019).
- Wang KH et al. (2018) Effects of collagen matrix and bioreactor cultivation on cartilage regeneration of a full-thickness critical-size knee joint cartilage defects with subchondral bone damage in a rabbit model. PLoS One 13: e0196779.
- Chen W et al. (2018) Autologous nasal chondrocytes delivered by injectable hydrogel for in vivo articular cartilage regeneration. Cell Tissue Bank 19: 35-46.
- Nixon AJ et al. (2017) Matrix-Induced autologous chondrocyte implantation (MACI) using a cell-seeded collagen membrane improves cartilage healing in the equine model. Journal of Bone and Joint Surgery 99: 1987-98.
- Zhao P et al. (2017) hWJECM-Derived Oriented Scaffolds with Autologous Chondrocytes for Rabbit Cartilage Defect Repairing. Tissue Eng Part A 24: 905-14.
- Minakuchi Y et al. (2004) Atelocollagen-mediated synthetic small interfering RNA delivery for effective gene silencing in vitro and in vivo. Nucleic Acids Res 32.
- Matsushita R et al. (2019) Repair of an Osteochondral Defect With Minced Cartilage Embedded in Atelocollagen Gel: A Rabbit Model. Am J Sports Med 47: 2216-24.
- Bellemère P (2018) Pyrocarbon implants for the hand and wrist. Hand Surg Rehabil 37: 129-54.
- Hannoun A et al. (2019) Pyrocarbon versus cobaltchromium in the context of spherical interposition implants: An in vitro study on cultured Chondrocytes. Eur Cell Mater 37: 1-15.
- Green B, Griffiths E, Almond S (2017) Neuropsychiatric symptoms following metal-on-metal implant failure with cobalt and chromium toxicity. BMC Psychiatry 17: 33.
FIGURE 1
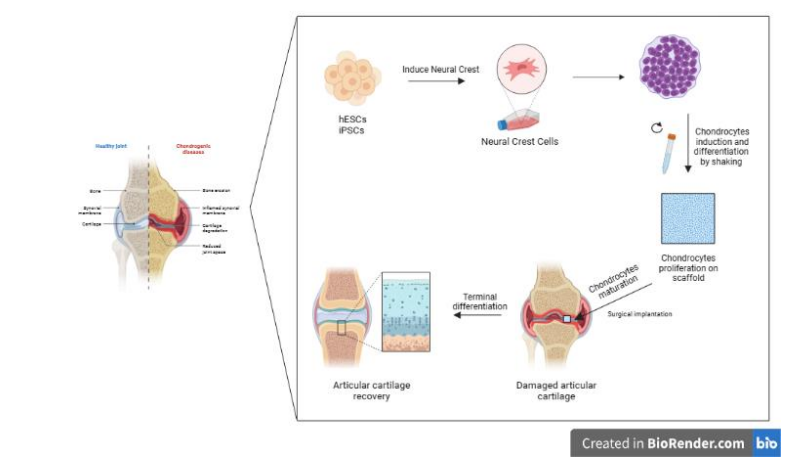
Figure 1: Chondrocytes derived-neural crest cells. Neural crest cells differentiation to increase chondrocytes. The use of scaffolds could improve site-specific chondrocytes incorporation to complete their maturation and promote the articular cartilage repair
Tables at a glance
Figures at a glance