Ex-Vivo Expansion of Bone Marrow-Derived Mesenchymal Stromal Cells forClinical use: The Starting Platelet Concentration of Human Platelet Lysate Affects Cell Proliferation, Senescence and Phenotype
Received Date: November 13, 2023 Accepted Date: December 13, 2023 Published Date: December 16, 2023
doi: 10.17303/jscr.2023.5.103
Citation: V. Becherucci, F. Nistico, L. Piccini, R. Ceccantini, F. Brugnolo et al. (2023) Ex-Vivo Expansion of Bone Marrow-Derived Mesenchymal Stromal Cells for Clinical use: The Starting Platelet Concentration of Human Platelet Lysate Affects Cell Proliferation, Senescence and Phenotype. J Stem Cell Rep 5: 1-23
Abstract
Background: In the last decades the replacement of fetal bovine serum (FBS) with human Platelet Lysate (hPL) for ATMPs expansion has been for a long time investigated to overcome FBS-related issues. Despite several studies confirming hPL safety and efficacy in Mesenchymal Stromal Cell (MSC) expansion, there are still gaps in the knowledge of hPL as a supplement, like the composition and release criteria. As growth factors are released after thrombocytes lysis during hPL production, starting platelet concentration may affect hPL quality. This study aimed to investigate hPL starting platelet concentration effects on bone marrow-derived MSC (BM-MSC) ex-vivo expansion.
Methods: MSC were isolated from the bone marrow (BM) of 7 donors and cultured from passage 1 to 5 in 4 different conditions: DMEM 10% FBS and DMEM 5% hPL varying starting platelet concentration. Particularly hPL was produced by in- -hospital Transfusion Service, in three different starting platelet concentrations (sPLTC): high (4x109 PLTS/ml), medium(2x109 PLTS/ml) and low (1x109 PLTS/ml). The study focused on the analysis of parameters that are mostly affected by hPL such as cell proliferation, immunophenotype, telomeric length, differentiation and senescence.
Results: Evaluation of proliferation indexes (PDT and PD) underlined dose-dependent effects of sPLTC, also confirmed by flow cytometry cell cycle analysis. Immunophenotype seems not to be affected by sPLTC. Differences were instead detected by adhesion molecules markers CD10, CD106, CD166, and CD146 expression, as their expression showed a dose-dependent downregulation, based on the sPLTC. Differentiation potential seems to be unaffected by different sPLTC as all cell batches differentiated into osteoblasts, adipocytes and chondrocytes. On the contrary senescence and relative telomeric length RTL (detected by SA-β-GAL activity and PNA-FITC flow cytometry) are strongly affected by sPLTC, in a dose-dependent manner. Particularly high sPLTC results in cell senescence associated with decreased RTL.
Conclusions: our data showed that sPLTC affects some BM MSCs properties, underlying its importance during hPL preparation. According to this study, we suggest a medium sPLTC for hPL preparation, as the best compromise between the increase in proliferation index and effects on senescence.
Keywords: Mesenchymal Stromal Cell; Human Platelet Lysate; Advanced Therapy Medicinal Product, Good Manufacturing Practices; Clinical Grade expansion
List of Abbreviations
MSCs: Mesenchymal stromal cells; ATMPs: advanced therapy medicinal products; GMP: Good Manufacturing Practices; EMA: European Medicines Agency; FBS: fetal bovine serum; hPL: human platelet lysate; TGF-β: transforming growth factor-beta; IGF-1: insulin-like growth factor-1; VEGF: vascular endothelial growth factor; FGF: fibroblast growth factor; PDGF: platelet-derived growth factor; EGF: epidermal growth factor; PLT: platelets; sPLTC: starting platelet concentration; BMNCs: Bone marrow mononuclear cells
Background
Mesenchymal stromal cells (MSCs) are self-renewing, multipotent progenitor cells with unique biologic properties that make them ideal candidates for cellular therapies. In addition to their ease of isolation and ex vivo expansion, MSCs possess peculiar characteristics that make them attractive for the treatment of various diseases [1]. First, MSCs can differentiate across various lineages beyond the conventional mesodermal lineages: thanks to this multipotency, MSCs found applications in regenerative medicine and tissue engineering [2].
Several studies have indicated that MSCs can provide therapeutic benefits through the secretion of soluble factors to induce an immunomodulatory environment [3], finding application in immunomodulation therapies [4-6], including the use of MSCs for COVID-19 pandemic disease [7,8].
Moreover, MSCs can migrate to sites of injury and tumour microenvironments: this unique tropism has allowed MSCs to serve as delivery vehicles for targeted therapy [9,10]. According to the definition given by the ISCT [11,12], MSCs are plastic-adherent when maintained in vitro and must be able to differentiate into osteoblasts, adipocytes and chondroblasts following standard cell culture differentiating conditions. In addition, ≥95% of the MSC population must express CD73, CD90 and CD105 and must lack expression of hematopoietic markers such as CD14, CD34, CD45 and HLA-DR [13]. Due to the low content of primary MSC in the bone marrow 0. 001–0.01% of total nucleated cells [14] significant ex vivo cell amplification before a clinical application is required to obtain therapeutic doses of 1–5 million cells/kg body weight [15,16]. The growing interest in the development of therapies based on MSCs is also evidenced by the presence of more than 150 clinical trials at www.clinicaltrials.gov (search term ‘Mesenchymal Stromal Cells’ accessed on 15 March 2023) This data is supported by the safety and feasibility of MSC-based therapies as no particular adverse effects and infusional toxicity have been observed [17]. To date, MSCs are considered advanced therapy medicinal products (ATMPs) by the European Medicines Agency (EMA) according to regulation No. [EC] 1394/2007 of the European Commission and must be produced in compliance with Good Manufacturing Practices (GMP) [18]. To address all these normative issues, overcoming the use of xenogeneic additives such as fetal bovine serum (FBS) in the clinical context is mandatory to avoid many risks, such as prion and viral transmission or adverse immunological reactions against xenogeneic components. Although in use for more than 50 years for culturing cells, FBS for clinical use presents many disadvantages coming from different points of view. For example, the FBS has never been fully characterized since serum in general is an ill-defined mixture of components in culture media, with qualitative and quantitative, geographical and seasonal batch-to-batch variations. Then the biosafety aspects since FBS may contain adverse factors, like endotoxin, mycoplasma, viral contaminants or prion proteins. Finally both ethical perspectives in terms of animal protection arguments regarding the harvest and collection of FBS from bovine fetuses and terms of recent concerns about the global supply versus demand of FBS [19].
Preliminary studies indicate that, in the last decades, the replacement of fetal bovine serum (FBS) with human Platelet lysate (hPL) is safe and efficient for MSC expansion. As a matter of fact, FBS has been studied for decades as a source of molecules and growth factors supporting MSC culture, without affecting cells' main features such as immunophenotype, immunomodulatory potential, and differentiation potential instead reporting a higher efficiency for MSC expansion [20-23]. While there are several examples in the literature of the use of platelet lysate for MSC clinical expansion [24-28], the debate is still open on hPL production, as it has been shown that several factors can influence hPL final quality, starting from the source of platelets concentrates [29], donor variability and manufacturing process [30-32]. A critical factor to consider in hPL production is the number of starting platelets since it affects the concentration of the growth factors released in the hPL such as transforming growth factor-beta (TGF-β), insulin-- like growth factor-1 (IGF-1), vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), plateletderived growth factor (PDGF) and epidermal growth factor (EGF) [33]. According with these considerations, our previous preliminary study on the different platelet concentrations has shown some interesting data, laying the background for further investigation [34]. A systematic meta-a-nalysis on the use of hPL for MSC culture underlined the benefits of using hPL instead of FBS but at the same time emphasizes the extreme heterogeneity in production methods suggesting the need for standardization, as small variations in the qualitative and quantitative composition of hPL, can determine major effects on cultured cells [35]. Recently scientific organizations and national entities, such as the AABB-ISCT Joint Working Group (American Association of Blood Banks and the International Society for Cell and Gene Therapy), the European Directorate for the Quality of Medicines & HealthCare of the Council of Europe (EDQM), and the International Society of Blood Transfusion (ISBT), have released criteria for the harmonization of raw materials such as hPL, giving indications about quality control and specifications for hPL production [36-41]. Despite these attempts, some aspects concerning platelet lysate remain to be clarified. For example, the concentration of specific growth factors as it is still not known which ones are required for optimum expansion [42] and this aspect is strictly connected to starting platelets concentration. According to guidelines [40], the minimum platelet content should be ≥ 2x1011unit but no further indications are provided, even if several studies reported that thrombocyte concentration is directly related to growth factor concentration/unit and may affect pro-proliferative effect [43,44]. The interesting study by Lange et al [44] underlined the importance of platelet concentration in hPL as a concentration below 1.5x106 PLT/µl significantly reduced the pro-proliferative effect. According to our experience [45], platelet concentrations must be in the range of 1.6 and 2.4 x106 PLT/µl to ensure an acceptable MSC expansion and we found [45] a moderate correlation between the number of platelets in each hPL and MSC population doublings as Spearman’s rank correlation coefficient at passage 1 was 0.7564, at passage 2 = 0.6543, and at passage 3 = 0.6764. This work aims to investigate how the starting platelets concentration may affect MSCs features.
Methods
hPL Production & Analysis
hPL Production
hPL was produced as previously described by our group [45]. Three hPL batches were prepared with different starting platelet concentrations (sPLTC). In comparison to our standardized protocol, where the platelet concentration is between 1.5-2.4x109 PLT/ml, for this study a high concentration platelet lysate (high sPLTC= 4x109PLT/ml) was produced following two dilutions with the same thawed freshfrozen AB group plasma to obtain a medium (2x109 PLT /ml) and low (1x109 PLT /ml) sPLTC concentration.
hPL analysis
An aliquot of hPL was collected for biochemical and growth factor analysis. Platelets were counted on Sysmex XN-1000™ Hematology Analyzer (Sysmex Europe GmbH), determination of total protein was performed by biuret-modified method on Dimension Vista® 1500 (Siemens Healthcare Diagnostics) and fibrinogen concentration was measured on Cobast 511® coagulation analyzer (Roche Diagnostics) with Clauss fibrinogen assay. Evaluation of concentrations levels of Platelet-Derived Growth Factor AB (PDGF-AB), Transforming Growth Factor β (TGFβ), Epidermal growth factor (EGF), Vascular-Endothelial Growth Factor (VEGF) and basic Fibroblast Growth Factor (bFGF) was performed using an ELISA kit (Invitrogen ELISA kit and Quantikine ELISA KIT Biotechne ), following manufacturer’s instructions.
BM-MSC Cultures and Analysis
BM-MSCs Production
Bone marrow mononuclear cells (BMNCs) were harvested from the iliac crest of 7 donors (mean age = 18) and equally split into 4 culture conditions at a seeding density of 100x106 BMNC/cm2 as follows:
- CONTROL Dulbecco's Modified Eagle Medium high glucose (D-MEM) (Invitrogen, Thermofisher) containing 10% of FBS (BioWest)
- D-MEM containing 5% high sPLTC (4x109PLT/ml) hPL
- D-MEM containing 5% medium sPLTC (2x109PLT/ml) hPL
- D-MEM containing 5% low sPLTC (1x109PLT/ml) hPL
After about 5 days, non-adherent cells were removed by two washes with saline (Baxter) and the adherent cells were refed every 3 days with the culture medium. Cells were detached at 85% confluence by adding TrypLE™ Select 10X (Thermofisher Scientific), microscopically counted with Bürker’s chamber and plated until passage 5 (p5) at 5.000 cells/cm2 in cell culture flasks (Corning ® Rectangular Canted Neck Cell Culture Flask with Vent Cap).
Evaluation of Cell Growth Kinetics
To evaluate the effects of different hPL on cellular growth, cumulative population doubling cPD and population doubling time PDT were calculated at each passage (from p1 to p5) using these equations:
- ● cPD=[log(n)-log(n0)]/log2
- ● PDT=time of culture (days)x[log(n)-log(n0)]/log2
where “n” is the number of cells detached and “n0” is the number of cells seeded. All counts were performed in triplicate. Cell cultures under different conditions were also observed with an inverted microscope at each passage (from p1 to p5) starting from the early time of culture (48 hours after seeding). A proliferation assay was performed by immunofluorescent staining with the incorporation of bromodeoxyuridine (BrdU) on BD FACS Canto I. Experiments were performed by using FITC BrdU Flow Kit (BD Pharmingen™) at passage 2. Cells were labelled for 72 hours and then analyzed. An aliquot of unlabeled cells was used as a negative control. Labelled and unlabeled cells were detached with TrypLE™ Select 10X and then fixed and permeabilized with BD Cytofix/Cytoperm Buffer following the manufacturer's instructions. Thereafter, cells were treated with DNase (30 µg of DNase/106 cells provided by the kit) to expose incorporated BrdU, incubated for 1 hour at 37°C washed 2 times and incubated for 15 minutes at room temperature with diluted fluorescent FITC anti-BrdU antibody and 7-AAD solution (both provided by the kit) for total DNA staining. Cells were acquired on the flow cytometer FACS Canto I (Becton Dickinson).
Immunophenotypic Analysis
To assess the possible effects of different culture conditions on the cell phenotype, a multiparameter flow cytometry analysis was set up at passages p1 to p5. Cells were washed with Dulbecco’s Phosphate buffered saline (DPBS 1x without Ca++/Mg++, Euroclone) and incubated for 15 minutes at room temperature with the following monoclonal antibodies cocktails:
- ● CD90 FITC (clone 5E10 cat.555595), CD105 PE (clone 266 cat.560839), CD73 PE (clone AD2 cat.550257), HLA-DR PE-Cy7(clone L243 cat.335830), CD45 APC-Cy7(clone 2D1 cat.348815), CD14 APC-Cy7(clone MΦp9 cat.333951), CD19 APC(clone SJ25C1 cat.345791), CD34 APC(clone 8G12 cat.345804) (BD biosciences) as characterized positive/negative surface MSC markers suggested by ISCT [46].
- ● CD10 FITC (clone HI10a cat.332775), CD166 PE(clone 3A6 cat.559263), CD146 PECy7(clone P1H12 cat. 562135), CD29 APC (clone MAR4 cat. 559883), CD106 PE (clone 51-10C9 cat. 555647), CD44 PE-Cy7 (clone G44-26 cat. 560533) and CD71 APC (clone M-A712 cat. 551374) (BD biosciences) as adhesion markers mostly affected by different concentrations of growth factors released in hPL [47].
- ● isotype controls APC-Cy7 IGg1 κ (clone MOPC-21 cat. 557873), PE-Cy7 IgG2a κ (clone G155-178 cat. 557907) APC-Cy7 Mouse IgG2bκ (clone 27-35 cat. 558061) APC IgG1κ (clone X40 cat. 567155), PE-Cy7 Mouse IgG1κ (clone MOPC-21 cat. 557872), PE IgG1κ (clone MOPC-21 cat. 555749), FITC Mouse IgG1k Isotype control BD (clone MOPC-31C cat. 550616) (BD biosciences).
The 7-amino actinomycin D (7-AAD, BD biosciences) was added to each tube to discriminate live/dead cells.
After incubation, cells were washed, resuspended in DPBS and acquired by flow cytometry on FACS Canto I (Becton Dickinson) equipped with the DIVA software program.
Trilineage Differentiation Assays
MSCs at p5 were used for differentiation assays. Undifferentiated MSCs, cultured under standard cell culture conditions, were used as controls (3x).
Osteogenic Induction
MSCs derived from FBS and hPL cultures were washed with DPBS and plated at 3.000 MSC/cm2 in 24 multiwell flat bottom plates (Euro Clone) until 90% confluence. Cell culture medium was replaced by osteogenic medium (StemPro™ Adipogenesis Differentiation Kit Gibco TM , ThermoFisher Scientific) exchanged every 2–3 days for 21 days. Differentiation of MSCs to osteocytes was determined by Alizarin Red staining of calcium matrix (Alizarin Red S, Sigma-Aldrich) after cold 4% paraformaldehyde fixation (Diapath S.p.A).
Adipogenic Induction
MSCs derived from FBS and hPL cultures were washed with DPBS and plated at 3.000 MSC/cm2 in 24 multiwell flat bottom plates (Euro Clone) with adipogenic medium StemPro™ (Adipogenesis Differentiation Kit Gibco™, ThermoFisher) following the manufacturer’s instructions. The medium was changed every 3 days and plates were observed under an inverted microscope for the development of lipid droplets (Nikon Eclipse TS100). Adipogenic differentiation was confirmed by Oil Red staining (Oil Red O; Sigma-Aldrich) after 15 days of culture in the differentiation medium. Cultures were fixed with 4% formaldehyde solution for 15 min over ice and rinsed with 60% isopropanol. For the identification of lipid droplets an Oil Red O working solution in 60% isopropanol was added to the cultures for 20 min at room temperature.
Chondrogenic Induction
MSCs were washed with DPBS and plated at 16.000 MSC/µl in 24 multiwell flat bottom plates (Euro Clone) for setting up the micro-mass. After 3 hours of 37°C incubation, a specific chondrogenic medium (StemPro™ Chondrogenesis Differentiation Kit, GibcoTM , ThermoFisher Scientific) was added and changed every 2–3 days for 15 days. Chondrogenic differentiation was confirmed by Alcian Blue staining of glycosaminoglycans (Alcian Blue 8GX, Sigma-Aldrich) after over-ice 4% paraformaldehyde fixation, following the manufacturer’s instructions.
Evaluation of Senescence
Another parameter that has been studied is the assessment of senescence as a phenomenon that can occur in cultured cells and lead to changes that make them ineffective for clinical use [48,49]. As previously analyzed in our last study [45], human platelet lysate does not seem to have these effects, but it is still interesting to investigate how and eventually if different platelet concentrations can induce early senescence phenomena.
Cells were morphologically evaluated under a 10X microscope (Nikon Eclipse TS100) to assess changes in size and morphology. Cell senescence was further investigated at passage 5 using two different methods: expression of beta-galactosidase activity (SA-β-Gal,) and detection of telomere length [50] in flow cytometry.
Evaluation of Senescence with Colourimetric Method
The senescence detection kit I (Promokine) was used, following the manufacturer’s instructions. Senescent cells display an increase in cell size and expression of the biochemical marker β galactosidase activity. After the removal of the culture medium and DPBS washing, cells were fixed for 15 minutes with a solution provided by the kit at room temperature. After paraformaldehyde fixation, cells were washed with DPBS (2x) and then fixed in a solution containing X-Gal. Cultures were incubated overnight at 37°C and observed under an optical microscope (Nikon Eclipse TS100) for the development of blue colour. To calculate the senescence index (SI), at least 100 cells were counted in triplicate. The result is expressed as the percentage of senescent cells as follows: SI=[(a)x100]/(a+b) where (a) is the average number of senescent cells and (b) the average number of non-senescent cells.
Evaluation of Senescence with Flow Cytometry
For the evaluation of senescence with flow cytometry, the Telomere PNA Kit/FITC for Flow Cytometry (Dako Cytomation Denmark) was used. Evaluation of senescence with relative telomere length (RTL) in flow cytometry is a method used to measure both the replicative history and senescence status of cells by determining the length of telomeres. A human erythromyeloblastoid leukaemia cell line (K562, provided by Laboratory of Cellular Manipulation, Hospital of Modena and Reggio Emilia, Italy) with known telomere length and DNA index was used as control. Briefly, MSCs at passage 5 were detached as previously described under section 3.1 and mixed in equal numbers with K562 cells. The cell suspension was exposed to DNA denaturation for 10 minutes at 82 °C in a microcentrifuge tube in presence of a hybridization solution without a probe or in presence of a fluorescein-conjugated PNA telomere probe. Overnight hybridization was done at room temperature in the dark followed by two 10-minute post-hybridization washes at 40 °C with a Wash Solution provided by the kit. Before flow cytometry analysis, an incubation with Propidium iodide (Propidium Iodide Staining Solution, BD Biosciences) was performed. Cells were analyzed by flow cytometry on a FACS Canto I. The RTL value is calculated as the ratio between the telomere signal of each sample and the control cell (K562 cell line) with correction for the DNA index of G0/1 cells by the following formula:
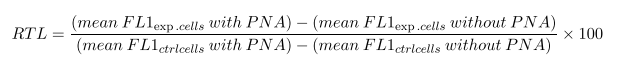
Statistical Analysis
Statistical analysis was performed with STATPLUS LE 7.6.5 software (AnalystSoft). Data are reported as mean± SD. Statistical differences were calculated with Analysis of Variance (ANOVA test) and T-test for paired data. Differences were considered significant at P-values< 0.05.
Results
Effects of different starting platelets concentration lysates on MSCs: Morphology and Proliferation
BM-MSC showed the characteristic fibroblast-like appearance, but those expanded in the presence of PL appeared smaller compared to those grown in FBS which are more spindle-shaped, elongated and showed denser cell bodies and cuboidal aspect. This characteristic is mainly observed in cells grown in DMEM with 5% high sPLTC platelet lysate (figure 1).
In flow cytometry analysis of physical parameters has confirmed morphological observation as MSCs grown in platelet lysate, especially at high sPLTC concentration, are smaller, less complex and more homogeneous compared to MSCs grown in FBS. This data is further confirmed by statistical analysis, performed with ANOVA test and t-test for paired data (FBS vs high sPLTC concentration platelet lysate), as shown in chart 1.
This data is further confirmed by flow cytometry analysis of the cell cycle with BrdU. As shown in chart n 2, the cells cultured in high and medium sPLTC concentration hPL show an increase in the proliferative phase than in other culture conditions. The data is statistically significant with p= 0.0002 calculated with one way ANOVA test at passage 2 ( cells have been analyzed after 72 hours from BrdU labelling).
The analysis of PD and PDT also supported this data. It has been observed that cells grown in platelet lysate at high and medium sPLTC concentrations have significantly higher PD. The data is marked in the early stages of cell culture (p2-p3), as confirmed by morphology and flow cytometric analysis of the cell cycle (chart n 3).
The PDT analysis confirmed these results. As shown in chart n 4, cells grown in FBS and low sPLTC hPL have a longer PDT compared to cells grown in medium sPLTC and high sPLTC platelet lysate. The effect has a progressive trend over time up to passage 5, showing a double PDT for cells grown in FBS and low sPLTC hPL.
Effects of different starting platelets concentration lysates on MSCs immunophenotype
The MSCs were analyzed at all culture passages, from p1 to p5, under 4 different culture conditions to verify the possible effects of different platelet concentrations on the cells' immunophenotype. All MSCs complied with the ISCT criteria [46]. As shown in Table 1, no significant differences were found regarding the expression of CD90, CD73, CD105, CD29, CD71, and CD44. Significant differences (p< 0.05) were found for CD10, CD106, CD166, and CD146 markers, (Figure 5). The percentage of expression of these markers shows a dose-dependent trend, based on the concentration of platelets used for the preparation of the lysate. The medium sPLTC and high sPLTC platelet lysate conditions show a roughly halved percentage of these markers compared to low sPLTC platelet lysate and FBS conditions, which however exhibit similar behaviour as already found for other analyzed parameters. Regarding, instead, the typically negative MSC markers such as CD45, CD34, and CD14, no increase in the percentage of expression in all the conditions analysed was detected (data not shown).
Effects of different starting platelets concentration lysates on trilineage differentiation
As shown in images 3 to 5 no differences were found between the four conditions and all MSCs differentiate in all mesodermal lineages (representative cases are reported).
Effects of different starting platelets concentration lysates on senescence
The high sPLTC hPL condition is associated with a significant increase of the senescent cells as shown in figure 6 (representative case) and in chart n 6, which shows the senescence index. This data correlates further with the RTL. As shown in chart 7, in high sPLTC hPL condition RTL is significantly lower than in other culture conditions.
Biochemical characterization of hPL
All platelet lysate aliquots used in this study were biochemically characterized. The results are shown in table n 2 where the concentration of the dosed growth factors PDGF-AB, TGFβ, EGF, VEGF and bFGF is proportional to the number of platelets used for the hPL preparation. However, for the plasma component (total protein and fibrinogen), no significant differences were found.
Discussion
Mesenchymal stromal cells are spindle-shaped plastic-adherent cells isolated from different tissue with multipotent differentiation capacity in vitro. Because of their biological characteristics, these cells currently represent the most frequently used cell type in advanced therapies with different purposes [1,2,4,5,51]. Overall, MSCs have shown promise as a cell therapy product, and numerous clinical trials are currently underway to investigate their safety and efficacy in a variety of conditions [17]. Since MSCs belong to the ATMPs category, they must be produced in compliance with GMP and all animal-derived culture supplements, such as FBS, should not be used for clinical grade expansion [52,18]. For decades, Hpl has been considered an excellent substitute for FBS for ATMPs expansion but despite extensive scientific evidence in terms of safety and efficacy [24-26,29,20], its production has not yet reached standardization. The recently introduced guidelines [36-41] represent a strong attempt to standardize and harmonize its production, but some aspects remain to be clarified.
In particular, the starting platelets concentration used for the hPL production is still a matter of discussion. The suggested minimum concentration is 2x1011/unit [40] but no further indications are provided, despite the evidence in the literature that platelet number affects the concentration of released growth factors and therefore efficacy in supporting cell expansion [31,33] According to an observation from our previous study [45] as a moderate correlation between the population doublings, and the number of platelets used was observed, this study aims to investigate if the starting platelet concentration used for hPL production, may affect some expanded MSC features. Based on obtained data it can be stated that the cell growth and the cell growth kinetics are affected by hPL starting platelets concentration. MSCs cultured in medium sPLTC and high sPLTC platelet lysate, show significantly higher PD than FBS and low sPLTC conditions. This data is also reflected in tandem with a halving of the PDT. The increased proliferative boost given by lysates with higher levels of platelets is particularly evident in the early stages of culture (p1-p2), as underlined by both microscopic and cell cycle analysis. An increase in cell proliferation can be observed after 72 hours of culture as well as at passage 2, just after 72 hours of culture, the percentage of cells in the S phase is significantly higher for MSCs cultured in medium sPLTC and high sPLTC platelet lysate than other conditions. The platelet lysate and the concentration of platelets used for its production, do not affect the expression of the classic ISCT surface markers CD105, CD90 and CD73. At the same time, typically negative markers such as CD45, CD34 and CD14 do not undergo any change in the percentage expression that remains below 2% of positivity, according to the ISCT criteria. This data further demonstrates how the use of platelet lysate represents a valid alternative to FBS for the GMP culture grade of mesenchymal stromal cells. Platelet lysate at any strating platelets concentration, does not alter the expression of markers that typically belong to the hematopoietic line and that allow to define the purity and quality of the MSC for clinical use [53].
Interestingly, the percentage of expression of CD166 (activated leukocyte cell adhesion molecule ALCAM), CD10 also known as neprilysin, CD106 (Vascular Cell Adhesion Molecule-1 VCAM-1) and CD146 (melanoma cell adhesion molecule MCAM) was significantly reduced in MCS grown in the presence of medium sPLTC and high sPLTC platelet lysate, in a dose-dependent manner. The CD166, a type-I transmembrane protein, belonging to the immunoglobulin superfamily, is described as an MSC marker expressed at higher levels in both Bone Marrow derived MSCs and Umbilical Cord MSCs [54]. Although its role in MSCs seems to be undetermined [55], cells expressing CD166 on their membrane are reported to have favourable chondrogenic differentiation capacity [56]. In our case, despite the different expression of CD166 no significative differences were found according to chondrogenic differentiation potential between the culture conditions.
Likewise, CD10 expression exhibits similar behaviour. This marker can be found on BM-MSC [57], but its expression seems to be unrelated to any particular MSC features. Moreover, the presence of CD10 on the MSC surface is still an object of discussion as described as positive in some papers and negative in other ones [58,59]. Even if the CD10 is downregulated in a dose-dependent manner according to the number of platelets used for the hPL preparation, seems not to modify the MSC features.
CD146 and CD106 BM-MSC surface markers are associated with clonogenicity, differentiation and immunomodulatory proprieties [60] and their expression decreases when MSC are differentiated into the osteogenic lineage, indicating their relevance as markers of MSC potency [61,62]. Moreover, the expression of CD146 has also been reported as associated with a higher immunomodulatory and secretory capacity, and thus therapeutic potency [63]. Our study shows that the starting platelet content of hPL affects their expression in a dose-dependent manner with progressive downregulation.
No differences were found between the culture conditions on the differentiation potential. Moreover,CD146 and CD106 are also associated with the MSC immunomodulatory potential. The CD106, a vascular adhesion protein and member of the immunoglobulin superfamily, binds to its target, the Integrin Subunit Alpha 4 (ITGA4), and recruits immune cells to the inflammatory site. It also has a role in MSC-mediated immunosuppression as the CD106 expression in MSCs can be induced by IFNγ treatment in presence of the pro-inflammatory cytokines tumour necrosis factor-alpha (TNF-α) or Interleukin-1-beta (IL-1β) [64]. The CD106 inhibition or gene knockout reduced MSC-induced immunosuppression in vitro and in vivo, indicating that CD106-mediated cell–cell contact is essential in immunoregulation by MSCs. This finding was supported in a study showing that CD106+ placental chorionic villi MSCs (CV-MSCs) better regulate helper T cells and produce higher levels of cytokines than CD106– CV-MSCs [65]. There are also reports indicating that the expression of CD146 is associated with better immune regulation. Upon inflammatory priming, CD146+ BM-MSCs had a greater secretory capacity, elevated expression of anti-inflammatory proteins, better immunosuppression of stimulated peripheral blood mononuclear cells (PBMCs) and T cells, and improved induction of regulatory T cells in vitro than the negative subpopulation [66]. Other studies reported that CD146 expression is downregulated during MSC prolonged culture and low expression levels are associated with in vitro and/or in vivo senescence and impaired migratory capacity [67-70]. These data are also confirmed in our study, where MSC grown in the presence of high sPLTC platelet lysate show greater signs of senescence. These phenomena are probably linked to the strong proliferative boost that the MSC grown in high sPLTC hPL receive, especially in the early stages of the culture.
Biochemical characterization of platelet lysate confirmed that the concentration of PDGF-AB, TGFβ, EGF, VEGF and bFGF was proportional to the number of platelets used for the hPL preparation. Interestingly, although platelet lysate contains a plethora of growth factors and cytokines, some growth factors appear to play a pivotal role in cell proliferation, survival and maintenance of the MSC differentiation/immunomodulation potential [71-73]. Transforming growth factor-beta (TGFβ) is known to influence cells from the chondrogenic lineage in vivo, promoting initial stages of mesenchymal condensation, prechondrocyte proliferation, production of extracellular matrix and cartilage-specific molecule deposition, while inhibiting terminal differentiation [74]. When applied to MSCs in vitro to study chondrocyte regeneration, cells show increased proliferation and a bias towards the chondrogenic lineage [75]. Along with TGFβ, PDGF-AB is the most powerful mitogenic agent for stimulating MSC's growth [76] and the most relevant growth factor in the human platelet lysate composition [77]. Many studies have deeply demonstrated its relevance in promoting and supporting MSC growth [44,78,79]. For example inhibition of PDGF-BB, bFGF and TGF-β by neutralizing antibodies reduced cell proliferation to a mean of 25% [71]. In our case the presence of a higher PDGF-AB and TGFβ concentration in high sPLTC platelet lysate explains the increased proliferative boost given to MSC in the early stages of culture, confirmed by proliferation index and cell cycle analysis. Moreover, even other growth factors such as EGF, VEGF and bFGF play an important role in supporting MSC proliferation, albeit in a lower concentration. Fibroblast growth factors (FGFs) are a family of growth factors involved in wound healing and angiogenesis. Among the various members of this family, FGF-2 or basic fibroblast growth factor (b-FGF) has been used in MSC-related studies showing increased rabbit, canine and human MSC proliferation in vitro [80]. The vascular endothelial growth factor increased MSC proliferation on its own even if MSCs do not express the VEGF receptor. This could imply that VEGF stimulates MSC proliferation by activation and downstream signalling of the platelet-derived growth factor (PDGF) receptors [81]. Finally, even the EGF enhances MSC self-renewal and expansion in vitro and has been found to promote MSC paracrine activity and osteogenic induction [82]. Although all these factors have been studied for a long time, the potential effects on the MSC's different features are not well characterised yet.
This research's strength lies in its thorough evaluation of cell growth and cycle analyses, which shed light on the complex relationship between sPLTC and MSC behavior. The main limitation of the study is represented by the relatively young donor group (average age of 18 years). Expanding the study to include older donors would undoubtedly represent an opportunity to enhance the clinical relevance of the findings, providing a more comprehensive understanding of how sPLTC may impact MSCs in patient populations that typically skew older. Overall we think that this research lays the groundwork for further exploration and potential applications in the field of regenerative medicine.
Conclusions
In conclusion, we suggest a starting platelet concentration for hPL preparation with a mean of 1,5-2,5x109 PLT/ml to guarantee efficient cell proliferation and avoid an excessive cell boost that could trigger MSC senescence.
Declarations
Not applicable
Consent for Publication
Not applicable
Availability of Data and Materials
All data generated or analysed during this study are included in this published article and its supplementary information files
Competing Interests
The authors declare that they have no competing interests
Funding
Not applicable
Acknowledgements
Not applicable
Authors' Contributions
VB, FN, LP and FB participated in the study concept and design, collection, analysis and interpretation of data. SE, EA, FB and GA participated in technical assistance and critical revision of the manuscript. LP, PP, BB, RC, EDR, VC and SB performed the platelet lysate preparation.VB, FB and FN performed the statistical analysis. VB and FN drafted the manuscript. All authors read and approved the final manuscript.
- Jovic D, Yu Y, Wang D, Wang K, Li H et al. (2022) A Brief Overview of Global Trends in MSC-Based Cell Therapy. Stem Cell Rev Rep 18: 1525-45.
- Capilla-Gonzalez V, Herranz-Perez V, Sarabia-Estrada R, Kadri N, Moll G (2022) Editorial: Mesenchymal Stromal Cell Therapy for Regenerative Medicine. Front Cell Neurosci 16: 932281.
- Song N, Scholtemeijer M, Shah K (2020) Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol Sci 41: 653-64.
- Wu X, Jiang J, Gu Z et al. (2020) Mesenchymal stromal cell therapies: immunomodulatory properties and clinical progress. Stem Cell Res Ther 11: 345.
- Jasim SA, Yumashev AV, Abdelbasset WK et al. (2022) Shining the light on clinical application of mesenchymal stem cell therapy in autoimmune diseases. Stem Cell Res Ther 13: 101.
- Kelly K, Rasko JEJ (2021) Mesenchymal Stromal Cells for the Treatment of Graft Versus Host Disease. Front Immunol. 12: 761616.
- Yao W, Shi L, Zhang Y et al. (2022) Mesenchymal stem/stromal cell therapy for COVID-19 pneumonia: potential mechanisms, current clinical evidence, and future perspectives. Stem Cell Res Ther 13: 124.
- Ciccocioppo R, Gibellini D, Astori G, Bernardi M, Bozza A et al. (2021) The immune modulatory effects of umbilical cord-derived mesenchymal stromal cells in severe COVID-19 pneumonia. Stem Cell Res Ther 12: 316.
- Zhang T, Lin R, Wu H, Jiang X, Gao J (2022) Mesenchymal stem cells: A living carrier for active tumor-targeted delivery. Adv Drug Deliv Rev 185: 114300.
- Hassanzadeh A, Altajer AH, Rahman HS, Saleh MM, Bokov DO et al. (2021) Mesenchymal Stem/Stromal Cell-Based Delivery: A Rapidly Evolving Strategy for Cancer Therapy. Front Cell Dev Biol 9: 686453.
- Viswanathan S, Shi Y, Galipeau J, Krampera M, Leblanc K et al. (2019) Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 21: 1019-24.
- Horwitz EM, Le Blanc K, Dominici M, Mueller I, Slaper-Cortenbach I et al. (2005) International Society for Cellular Therapy. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 7: 393-5.
- Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F et al. (2006) Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8: 315-7.
- Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R et al. (1999) Multilineage potential of adult human mesenchymal stem cells. Science 284: 143-7.
- Lee OJ, Luk F, Korevaar SS, Koch TG, Baan CC et al. (2020) The Importance of Dosing, Timing, and (in)Activation of Adipose Tissue-Derived Mesenchymal Stromal Cells on Their Immunomodulatory Effects. Stem Cells Dev 29: 38-48.
- Wang Z, Wang L, Su X, Pu J, Jiang M, He B (2017) Rational transplant timing and dose of mesenchymal stromal cells in patients with acute myocardial infarction: a meta-analysis of randomized controlled trials. Stem Cell Res Ther 8: 21.
- Wang Y, Yi H, Song Y (2021) The safety of MSC therapy over the past 15 years: a meta-analysis. Stem Cell Res Ther 12: 545.
- European Commission (2007) REGULATION (EC) No 1394/2007 of the European Parliament and of The Council of 13 November 2007 on advanced therapy medicinal products and amending Directive 2001/83/EC and Regulation (EC) No 726/2004.
- Gstraunthaler G, Lindl T, van der Valk JA (2013) A plea to reduce or replace fetal bovine serum in cell culture media. Cytotechnology 65: 791-3.
- Guiotto M, Raffoul W, Hart AM, Riehle MO, di Summa PG (2020) Human platelet lysate to substitute fetal bovine serum in hMSC expansion for translational applications: a systematic review. J Transl Med 18: 351.
- Doucet C, Ernou I, Zhang Y, Llense JR, Begot L, Holy X, et al. (2005) Platelet lysates promote mesenchymal stem cell expansion: a safety substitute for animal serum in cell-based therapy applications. J Cell Physiol 205: 228-36.
- Burnouf T, Strunk D, Koh MBC, Schallmoser K (2016) Human platelet lysate: replacing fetal bovine serum as a gold standard for human cell propagation? Biomaterials 76: 371-87.
- Fekete N, Rojewski MT, Furst D, Kreja L, Ignatius A et al. (2012) GMP-compliant isolation and large-scale expansion of bone marrow-derived MSC. PLoS One 7: e43255.
- Mareschi K, Marini E, Niclot AGSB, Barone M, Pinnetta G et al. (2022) A New Human Platelet Lysate for Mesenchymal Stem Cell Production Compliant with Good Manufacturing Practice Conditions.Int J Mol Sci 23: 3234.
- Oeller M, Laner-Plamberger S, Krisch L, Rohde E, Strunk D, Schallmoser K (2021) Human Platelet Lysate for Good Manufacturing Practice-Compliant Cell Production. International Journal of Molecular Sciences 22: 5178.
- Capelli C, Domenghini M, Borleri G, Bellavita P, Poma R et al. (2007) Human platelet lysate allows expansion and clinical grade production of mesenchymal stromal cells from small samples of bone marrow aspirates or marrow filter washouts. Bone Marrow Transplant 40: 785-91.
- Astori G, Bernardi M, Bozza A et al. (2020) Logistics of an advanced therapy medicinal product during COVID-19 pandemic in Italy: successful delivery of mesenchymal stromal cells in dry ice. J Transl Med 18: 451.
- Ciccocioppo R, Gibellini D, Astori G, Bernardi M, Bozza A et al. (2021)The immune modulatory effects of umbilical cord-derived mesenchymal stromal cells in severe COVID-19 pneumonia. Stem Cell Res Ther 12: 316.
- Schallmoser K et al. (2007) Human platelet lysate can replace fetal bovine serum for clinical-scale expansion of functional mesenchymal stromal cells. Transfusion 47: 1436-46.
- Fekete N, Gadelorge M, Furst D, Maurer C, Dausend J et al. (2012) Platelet lysate from whole blood-derived pooled platelet concentrates and apheresis-derived platelet concentrates for the isolation and expansion of human bone marrow mesenchymal stromal cells: production process, content and identification of active . components. Cytotherapy 14: 540-54.
- Bernardi M, Agostini F, Chieregato K, Amati E, Durante C et al. (2017) The production method affects the efficacy of platelet derivatives to expand mesenchymal stromal cells in vitro. J Transl Med 15: 90.
- Agostini F, Polesel J, Battiston M, Lombardi E, Zanolin S et al. (2017) Standardization of platelet releasate products for clinical applications in cell therapy: a mathematical approach. J Transl Med 15: 107.
- Viau S, Lagrange A, Chabrand L, Lorant J, Charrier M et al. (2019) A highly standardized and characterized human platelet lysate for efficient and reproducible expansion of human bone marrow mesenchymal stromal cells. Cytotherapy 21: 738-54.
- Becherucci V, Nistico F, Piccini L,Gori V et al. (2020) Platelet Lysate as source of growth factors for Bone Marrow derived Mesenchymal Stromal cells clinical expansion: a study of starting platelet concentration dose-dependent effects.Cytotherapy 22: 67-8.
- Palombella S, Perucca Orfei C, Castellini G et al. (2022) Systematic review and meta-analysis on the use of human platelet lysate for mesenchymal stem cell cultures: comparison with fetal bovine serum and considerations on the production protocol. Stem Cell Res Ther 13: 142.
- Strunk D, Lozano M, Marks DC, Loh YS, Gstraunthaler G et al. (2018) International Forum on GMP-grade human platelet lysate for cell propagation. Vox Sang 113: e1-25.
- Bieback K, Fernandez-Muñoz B, Pati S, Schäfer R (2019) Gaps in the knowledge of human platelet lysate as a cell culture supplement for cell therapy: a joint publication from the AABB and the International Society for Cell & Gene Therapy. Cytotherapy 21: 911-24.
- Henschler R, Gabriel C, Schallmoser K, Burnouf T, Koh MBC (2019) Human platelet lysate current standards and future developments. Transfusion 59: 1407-13.
- European Directorate for the Quality of Medicines & HealthCare of the Council of Europe [EDQM] (2020b). Raw Materials of Biological Origin for the Production of Cell-based and Gene Therapy Medicinal Products.European Pharmacopoeia, 10th Edn, Chapter 5.2.12, Council of Europe. Strasbourg: EDQM.
- Schallmoser K, Henschler R, Gabriel C, Koh MBC, Burnouf T (2020) Production and Quality Requirements of Human Platelet Lysate: A Position Statement from the Working Party on Cellular Therapies of the International Society of Blood Transfusion. Trends Biotechnol 38: 13-23.
- Chou M, Burnouf T (2017) Current methods to manufacture human platelet lysates for cell therapy and tissue engineering: possible trends in product safety and standardization. VOXS 12: 168-75.
- Shih DT, Burnouf T (2015) Preparation, quality criteria, and properties of human blood platelet lysate supplements for ex vivo stem cell expansion. N Biotechnol 32:199-211.
- Schallmoser K, Strunk D (2013) Generation of a pool of human platelet lysate and efficient use in cell culture. Methods Mol Biol 946: 349-62.
- Lange C, Cakiroglu F, Spiess AN, Cappallo-Obermann H, Dierlamm J, Zander AR (2007) Accelerated and safe expansion of human mesenchymal stromal cells in animal serum-free medium for transplantation and regenerative medicine. J Cell Physiol 213: 18-26.
- Becherucci V, Piccini L, Casamassima S, Bisin S, Gori V et al. (2018) Human platelet lysate in mesenchymal stromal cell expansion according to a GMP grade protocol: a cell factory experience. Stem Cell Res Ther 9: 124.
- Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F et al. (2006) Minimal criteria for defining multipotent mesenchymal stromal cells. The ISCT position statement.Cytotherapy. 8: 315-7.
- Reis M, McDonald D, Nicholson L, Godthardt K, Knobel S et al. (2018) Global phenotypic characterisation of human platelet lysate expanded MSCs by high-throughput flow cytometry. Sci Rep 8: 3907.
- Baxter MA, Wynn RF, Jowitt SN, Wraith JE, Fairbairn LJ, Bellantuono I (2004) Study of telomere length reveals rapid aging of human marrow stromal cells following in vitro expansion. Stem Cells 22: 675-82.
- Turinetto V, Vitale E, Giachino C (2016) Senescence in Human Mesenchymal Stem Cells: Functional Changes and Implications in Stem Cell-Based Therapy. Int J Mol Sci 17: 1164.
- Lauzon W, Sanchez Dardon J, Cameron DW, Badley AD (2000) Flow cytometric measurement of telomere length. Cytometer 42: 159-64.
- Zhao S, Wehner R, Bornhäuser M, Wassmuth R, Bachmann M, Schmitz M (2010) Immunomodulatory properties of mesenchymal stromal cells and their therapeutic consequences for immune-mediated disorders.Stem Cells Dev 19: 607-14.
- EMEA/CHMP Guideline on human cell-based medicinal products.EMEA Editor. London, UK: 7 Westferry Circus, Canary Wharf; 2008.
- Amati E, Sella S, Perbellini O, Alghisi A, Bernardi M et al. (2017) Generation of mesenchymal stromal cells from cord blood: evaluation of in vitro quality parameters prior to clinical use. Stem Cell Res Ther 8: 14.
- Brinkhof B, Zhang B, Cui Z, Ye H, Wang H (2020) ALCAM (CD166) as a gene expression marker for human mesenchymal stromal cell characterisation. Gene X 5: 100031.
- Moraes DA, Sibov TT, Pavon LF, Alvim PQ, Bonadio RS et al. (2016) A reduction in CD90 (THY-1) expression results in increased differentiation of mesenchymal stromal cells. Stem Cell Res Ther 7: 97.
- Jonitz A, Lochner K, Peters K, Salamon A, Pasold J, Mueller-Hilke B, Hansmann D, Bader R (2011) Differentiation capacity of human chondrocytes embedded in alginate matrix. Connect. Tissue Res 52: 503-11.
- Rojewski MT, Weber BM, Schrezenmeier H (2008) Phenotypic Characterization of Mesenchymal Stem Cells from Various Tissues. Transfus Med Hemother 35: 168-84.
- Mafi P, Hindocha S, Mafi R, Griffin M, Khan WS (2011) Adult mesenchymal stem cells and cell surface characterization - a systematic review of the literature. Open Orthop J 5: 253-60.
- Amati E, Perbellini O, Rotta G et al. (2018) High- -throughput immunophenotypic characterization of bone marrow- and cord blood-derived mesenchymal stromal cells reveals common and differentially expressed markers: identification of angiotensin-converting enzyme (CD143) as a marker differentially. expressed between adult and perinatal tissue sources. Stem Cell Res Ther 9: 10.
- Srinivasan A, Sathiyanathan P, Yin L, Liu TM, Lam A et al. (2022) Strategies to enhance immunomodulatory properties and reduce heterogeneity in mesenchymal stromal cells during ex vivo expansion. Cytotherapy 24: 456-72.
- Liu F, Akiyama Y, Tai S, Maruyama K, Kawaguchi Y, Muramatsu K, Yamaguchi K (2008) Changes in the expression of CD106, osteogenic genes, and transcription factors involved in the osteogenic differentiation of human bone marrow mesenchymal stem cells. J Bone Miner Metab 26: 312-20.
- Delorme B, Ringe J, Gallay N, Le Vern Y, Kerboeuf D et al. (2008) Specific plasma membrane protein phenotype of culture-amplified and native human bone marrow mesenchymal stem cells. Blood 111: 2631-5.
- Bowles AC, Kouroupis D, Willman MA, Perucca Orfei C, Agarwal A, Correa D (2020) Signature quality attributes of CD146+ mesenchymal stem/stromal cells correlate with high therapeutic and secretory potency. Stem Cells 38: 1034-49.
- Ren G, Zhao X, Zhang L, Zhang J, L'Huillier A et al. (2010) Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J Immunol 184: 2321-8.
- Yang ZX, Han ZB, Ji YR, Wang YW, Liang L et al. (2013) CD106 identifies a subpopulation of mesenchymal stem cells with unique immunomodulatory properties. PLoS One 8: e59354.
- Bowles AC, Kouroupis D, Willman MA, Perucca Orfei C, Agarwal A, Correa D (2020) Signature quality attributes of CD146+ mesenchymal stem/stromal cells correlate with high therapeutic and secretory potency. Stem Cells 38: 1034-49.
- Li Y, Wu Q, Wang Y, Li L, Bu H, Bao J (2017) Senescence of mesenchymal stem cells (Review). Int J Mol Med 39: 775-82.
- Kwon O, Jung E, Kwon KS, Cho YS, Rhee SK (2011) Evidences for correlation between the reduced VCAM-1 expression and hyaluronan synthesis during cellular senescence of human mesenchymal stem cells. Biochem Biophys Res Commun 404: 463-9.
- Weng Z, Wang Y, Ouchi T, Liu H, Qiao X et al. (2022) Mesenchymal Stem/Stromal Cell Senescence: Hallmarks, Mechanisms, and Combating Strategies. Stem Cells Transl Med 11: 356-71.
- Jin HJ, Kwon JH, Kim M, Bae YK, Choi SJ et al. (2016) Down regulation of Melanoma Cell Adhesion Molecule (MCAM/CD146) Accelerates Cellular Senescence in Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells. Stem Cells Transl Med 5: 427-39.
- Oeller M, Laner-Plamberger S, Krisch L, Rohde E, Strunk D, Schallmoser K (2021) Human Platelet Lysate for Good Manufacturing Practice-Compliant Cell Production. Int J Mol Sci 22: 5178.
- Jung S, Sen A, Rosenberg L, Behie LA (2010) Identification of growth and attachment factors for the serum-free isolation and expansion of human mesenchymal stromal cells. Cytotheraphy 12: 637-57.
- Fekete N, Gadelorge M, Fürst D, Maurer C, Dausend J et al. (2012) Platelet lysate from whole blood-derived pooled platelet concentrates and apheresis-derived platelet concentrates for the isolation and expansion of human bone marrow mesenchymal stromal cells: production process, content and identification of active components. Cytotherapy 14: 540-54.
- de Araújo Farias V, Carrillo-Gálvez AB, Martín F, Anderson P (2018) TGF-β and mesenchymal stromal cells in regenerative medicine, autoimmunity and cancer. Cytokine Growth Factor Rev 43: 25-37.
- Longobardi L, O'Rear L, Aakula S, Johnstone B, Shimer K (2006) Effect of IGF-I in the chondrogenesis of bone marrow mesenchymal stem cells in the presence or absence of TGF-beta signaling. J Bone Miner Res 21: 626-36.
- Kang YJ, Jeon ES, Song HY, Woo JS, Jung JS et al. (2005) Role of c-Jun N-terminal kinase in the PDGF-induced proliferation and migration of human adipose tissue-derived mesenchymal stem cells. J Cell Biochem 95: 1135-45.
- Burnouf T, Goubran HA, Chen TM, Ou KL, El-Ekiaby M, Radosevic M (2013) Blood-derived biomaterials and platelet growth factors in regenerative medicine. Blood Rev 27: 77-89.
- Astori G, Amati E, Bambi F, Bernardi M, Chieregato K et al. (2016) Platelet lysate as a substitute for animal serum for the ex-vivo expansion of mesenchymal stem/stromal cells:present and future.Stem Cell Res Ther 7: 93.
- Burnouf T, Strunk D, Koh MB, Schallmoser K (2016) Human platelet lysate: Replacing fetal bovine serum as a gold standard for human cell propagation? Biomaterials 76: 371-87.
- Ramasamy R, Tong CK, Yip WK, Vellasamy S, Tan BC, Seow HF (2012) Basic fibroblast growth factor modulates cell cycle of human umbilical cord-derived mesenchymal stem cells. Cell Prolif 45: 132-9.
- Ball SG, Shuttleworth CA, Kielty CM (2007) Mesenchymal stem cells and neovascularization: role of plateletderived growth factor receptors. J Cell Mol Med 11: 1012-30.
- Tamama K, Kawasaki H, Wells A (2010) Epidermal growth factor (EGF) treatment on multipotential stromal cells (MSCs). Possible enhancement of therapeutic potential of MSC. J Biomed Biotechnol 2010: 795385.
- Becherucci V, Gori V, Filì l et al. (2013) Fibroblast growth factor 2 exerts different unexpected effects on bone marrow-derived human mesenchymal stromal cells,Cytotherapy 15: S15-6,
FIGURE 1
FIGURE 2
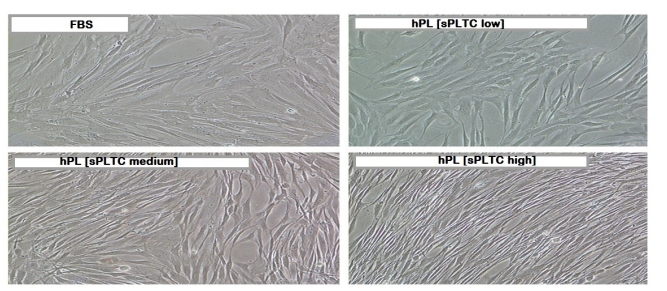
Figure 1: Representative microscope image of cells grown under four different conditions (magnification 10X). Cells grown in platelet lysate are smaller and elongated compared to cells grown in FBS. The effect is more pronounced for the high concentration hPL condition [high sPLTC (4x109sup> PLT/ml)]. FBS= Fetal Bovine Serum, hPL SPLTC = human platelet lysate starting platelets concentration.
FIGURE 3
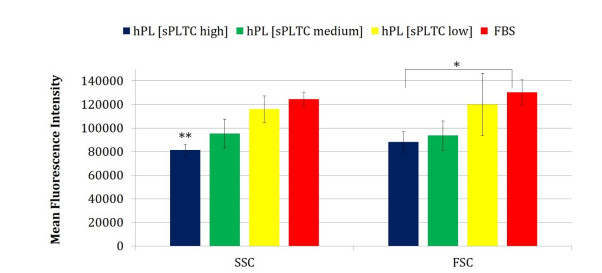
Chart 1: The chart shows the MFI (Mean Fluorescence Intensity) of the physical parameters FSC and SSC under 4 different culture conditions. Statistical analysis was conducted using ANOVA (Analysis of Variance) and significance was found for the FSC parameter with *p < 0.05 (=0.001). Additionally, a paired t-test was performed between the FBS and the high sPLTC platelet lysate condition, and significance was also found for the SSC parameter (significance with ** p < 0.05 = 0.001). Data are represented as mean ±SD (n=420)
FIGURE 4
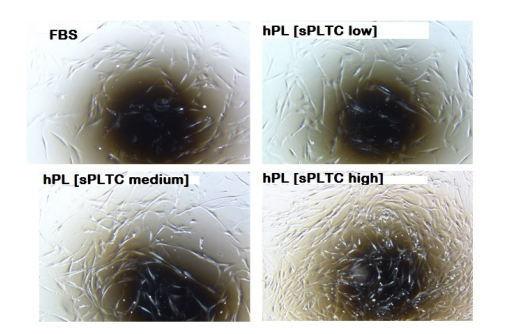
Figure 2: Representative microscope (4X) image of cells grown under 4 different conditions and observed after 72 hours of culture at passage 2. Cells grown in high sPLTC concentration hPL have a higher proliferative boost than other culture conditions. FBS= Fetal Bovine Serum, hPL SPLTC = human platelet lysate starting platelets concentration.
FIGURE 5
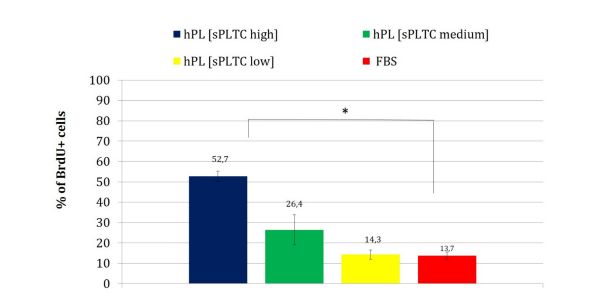
Chart 2: The chart presents the percentage of BrdU-positive cells (S-phase cells in active proliferation), after 72 hours of culture at passage 2.Data are represented as mean ±SD with n= 84. Statistical analysis was performed with one way ANOVA test (significance with * p < 0.05,p=0.0002). FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration.
FIGURE 6
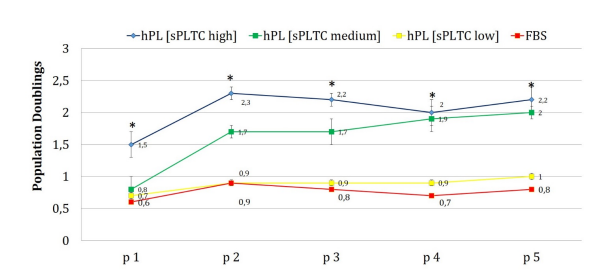
Chart 3: The chart represents the population doubling (PD) of MSCs grown in DMEM 5% platelet lysate at three different starting platelets concentration (high, medium, low) and MSCs grown in DMEM 10% FBS. Cells grown in platelet lysate, especially at high and medium starting platelets concentration, had a significantly higher proliferative potential, particularly at early passages. Cells cultured in FBS and low sPLTC lysate, follow a similar trend along the different culture passages. Data are represented as mean ±SD with n=420. Statistical analysis was performed with one way ANOVA test (significance with * p < 0.05, p=0.00001). FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration.
FIGURE 7
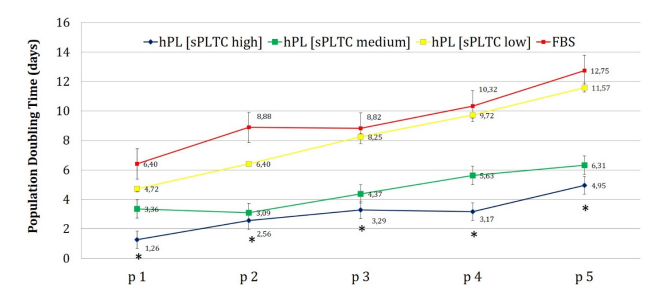
Chart 4: The chart represents the population doubling time (PDT) of MSCs grown in DMEM 5% platelet lysate at the three different platelets concentration and MSCs grown in DMEM 10% FBS. Cells grown in sPLTC and high sPLTC platelet lysate, from p1 to p5, take half time to double their population compared to cells grown in FBS and low sPLTC PL. This trend is most visible at early culture passages where cells grown in FBS take five times longer than cells grown in high sPLTC platelet lysate to double their population. Data are reported as mean ±SD n=420. Statistical analysis was performed with one way ANOVA test (significance with *p < 0.05, p=0.00001). FBS= Fetal Bovine Serum, hPL SPLTC = human platelet lysate starting platelets concentration.
FIGURE 8
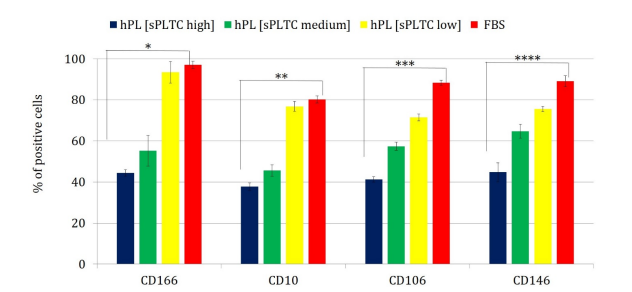
Chart 5: The chart shows the percentage of cells expressing the CD10, CD 106, CD 166 and CD146 markers in the 4 different culture conditions. Statistical analysis was performed with one way ANOVA test, and significance was found for all the analyzed parameters. Significance of the test with p < 0,05 (*p CD166 =0,004, **p CD10 =0,036, ***p CD106 = 0,001,****p CD146 =0,0024). Data are presented as mean ±SD n=420 (calculated from p1 to p5), FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration.
FIGURE 9
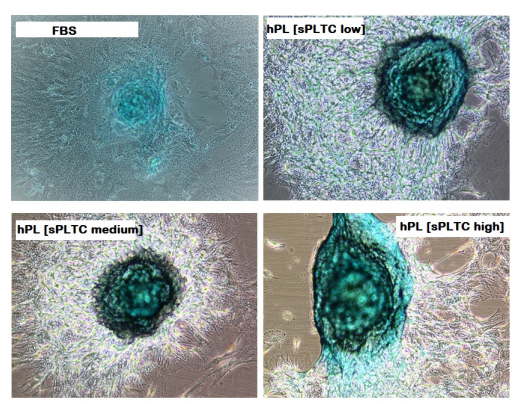
Figure 3: Representative case of MSC chondrogenic differentiation at p5 in 4 different culture conditions. Alcian blue staining shows hyaluronic acid for chondrocytes, after 15 days of induction. n= 84 FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration
FIGURE 10
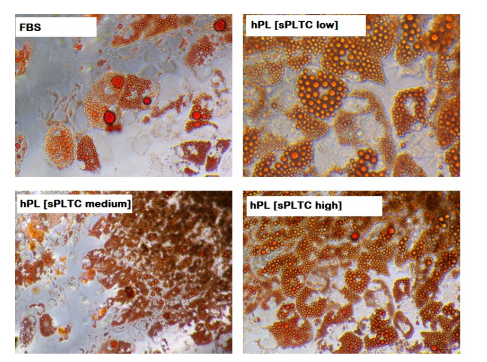
Figure 4: Representative case of MSC adipogenic differentiation at p5 in 4 different culture conditions. Oil Red O shows intracytoplasmatic vacuoles, after 15 days of adipogenic induction. n= 84 FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration
FIGURE 11
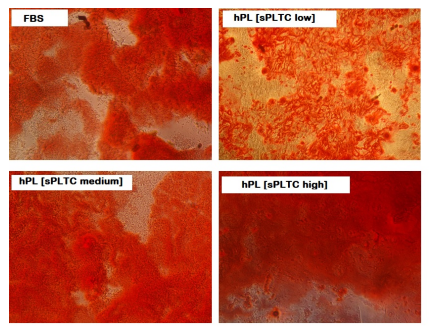
Figure 5: Representative case of MSC osteogenic differentiation at p5 in 4 different culture conditions. Alizarin Red S staining shows the presence of calcium matrix in osteoblasts, after 21 days of osteogenic induction. n= 84 FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration.
FIGURE 12
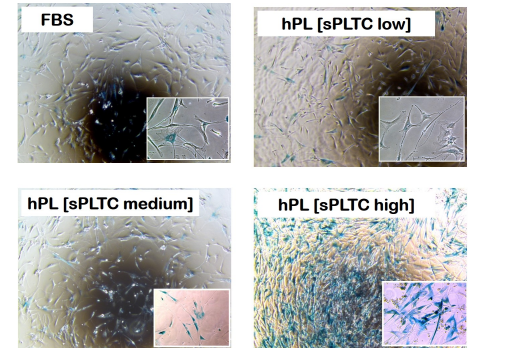
Figure 6: Representative case of histochemical detection of Senescence-associated marker β-galactosidase on MSC at passage 5, in 4 different culture conditions. n= 84 FBS= Fetal Bovine Serum, hPL sPLTC= human platelet lysate starting platelets concentration.
FIGURE 13
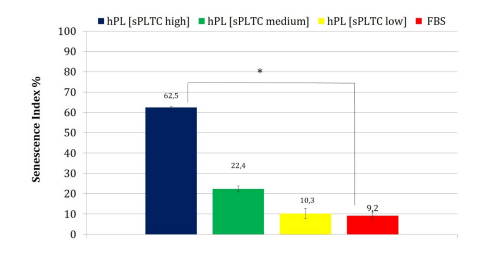
Chart 6: The chart shows the senescence index of MSC in the 4 different culture conditions at p5. The high sPLTC hPL condition shows a significant increase in senescent cells (one way ANOVA test with n= 84 and *p= 0.00001 Data are represented as mean ±SD FBS= Fetal Bovine Serum, hPL sPLTC = human platelet lysate starting platelets concentration.
FIGURE 14
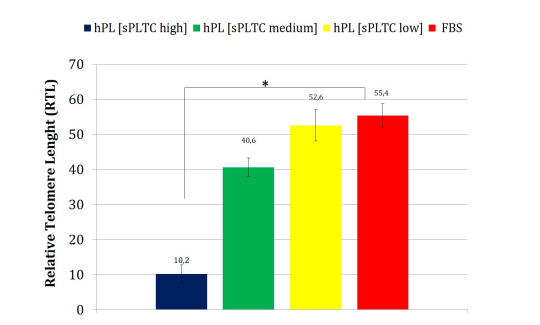
Chart 7: The chart shows the relative telomere length (RTL) of MSCs in the 4 different culture conditions at p5. The high sPLTC hPL condition shows a significant reduction of RTL (one way ANOVA test with n= 84 and *p= 0.00002 Data are represented as mean ±SD FBS= Fetal Bovine Serum, hPL SPLTC = human platelet lysate starting platelets concentration.
Tables at a glance
Figures at a glance