Glypican-1, -4 and 6 Participate in the Hair Follicle Stem Cell Differentiation viaSpecific Interactions with BMP2R, Fz7 and Patched 1
Received Date: September 05, 2024 Accepted Date: October 05, 2024 Published Date: October 08, 2024
doi: 10.17303/jscr.2024.6.103
Citation: Charlie Colin-Pierre, Laurent Ramont, Stéphane Brézillon (2024) Glypican-1, -4 and 6 Participate in the Hair Follicle Stem Cell Differentiation via Specific Interactions with BMP2R, Fz7 and Patched 1. J Stem Cell Rep 6: 1-15
Abstract
During hair shaft formation, the role of the human hair follicle dermal papilla cells (HHFDPC), the keratinocytes of outer root sheath (KORS) or the human dermal microvascular endothelial cells (HDMEC) was analyzed on the hair follicle stem cell (HFSC) fate regulation. The role of glypicans (GPCs) was investigated in the regulation of the HFSC differentiation. The effect of the conditioned media of HHFDPCs (HHFDPC CM), KORS (KORSCM) or HDMECs (HDMECCM) on HFSC differentiation was performed using specific biomarkers. The growth factors secreted by HHFDPC, KORS and HDMEC and the association of GPCs with their receptors were investigated. The role of the heparan sulfate-type glycosaminoglycan (GAG) chains was analyzed on the modulation of HFSC differentiation. HHFDPCs expressed Wnt and BMP and induced HFSC differentiation into secondary hair germ (SHG) progenitors. In contrast, KORS and HDMEC expressed Shh and BMP and induced HFSC differentiation into ORS progenitors. An association of GPC1, 4 and 6 with BMPR2, Fz7 and Patched 1 was demonstrated. The HFSC differentiation was altered by heparinase III. This study highlights the role of KORS and endothelial cells in the regulation of HFSC fate and the role of GPC1, 4 and 6 during HFSC differentiation.
Keywords: Hair Follicle; Stem Cell Differentiation; Glypicans; BMP; Wnt and Shh Signaling Pathways
Introduction
The hair follicle (HF) is a complex mini organ allowing the growth of new hair shafts. The deepest part of the HF is the bulb which is composed of the dermal papilla and the hair matrix. The dermal papilla is an invagination, in the hair matrix, of connective tissue essentially composed of papillary fibroblasts with a small capillary network in its center. Above the bulb, the HF is formed by concentric zones, from inside to outside: the hair shaft, the inner root sheath (IRS) and the outer root sheath (ORS) which contains the bulge, the reserve of hair follicle stem cell (HFSC) [1].
Hairs are constantly renewed, and forty to one hundred hairs are lost per day. The renewal of hair shafts involves cyclic changes in the HF [2,3]. Hair growth cycle is divided into three main phases: anagen (hair shaft growth), catagen (secondary hair germ (SHG) formation) and telogen (resting phase). The hair cycle is centered on the activation of pluripotent cells of the HF to induce differentiation and to provide different cell lineages of the HF [4].
Several hypotheses on the pattern of differentiation of these cells have been made over time [1,5], demonstrating the complexity of this process and the difficulty of studying it, especially in humans. Nevertheless, the cross-referencing of recent data on human hair multipotent cells with analyses performed in mice allows a better understanding of the lineage of these cells. In brief, Thve major distinct populations are defined in the literature according to their location, origin, differentiation during the phases of the cycle and their cell markers [5-9] (i) Bulge stem cells are able to differentiate in all the cell types forming the HF. They express cytokeratin 15 (cK15) and α6 integrin subunit (ITGA6) as well as OX-2 membrane glycoprotein (CD200); (ii) ORS progenitor cells differentiate to form ORS and participate in the formation of the SHG. They express CD34; (iii) SHG progenitor cells are formed under the bulb during the catagen phase and differentiate to form the matrix and the ORS. They express CD200 and Leucine-rich repeat-containing G-protein coupled receptor 5 (LGR5); (iv) Matrix transit-amplifying (TA) cells allow the IRS and hair shaft formations. They express lymphoid enhancer-binding factor 1 (Lef1). (v) Differentiated cells are represented by ORS, IRS and hair shaft cells.
The process of HF stem cell differentiation involves many cell types: keratinocytes (of the IRS or of the ORS), fibroblasts of the dermal papilla, endothelial cells (hair capillaries), adipose cells and immune system cells. The interaction between these different cell types makes the study of the hair growth cycle regulation complex. The intercellular communication is still poorly characterized but it is known that several signaling pathways and growth factors regulate the HFSC differentiation [10]. The bone morphogenetic protein (BMP) pathway plays an important role during the telogen phase. In particular, BMP2 and 4 secreted by the dermal papilla and adipose tissue, promote the HF stem cell quiescence [11,12]. In contrast, the Wnt signaling pathway allows the telogen to anagen transition. In particular, the secretion of platelet-derived growth factor subunit A (PDGFA) by the adipose tissue activates the Wnt pathway within the dermal papilla. In particular, sonic hedgehog (Shh), secreted by the matrix TA cells, regulates in the formation of the new pool of ORS progenitor cells necessary to the next growth cycle [13].
This brief introduction is far from reflecting the intense complexity of the crosstalk required for the regulation of the hair growth cycle and the hair shaft formation. This level of regulation involves numerous cell types and is allowed thanks to multiple growth factors and signaling pathways. Only few studies provide evidence of the implication of heparan sulfate proteoglycans (HSPGs) in these regulatory processes. Indeed, it has been shown that within the HF the distribution of various HSPGs varies during the hair growth cycle [14-19]. Moreover, HSPGs are known to regulate the cell proliferation, migration, and differentiation induced by growth factors in several organs and tissues [20,21]. In particular, glypicans (GPCs), membrane HSPGs composed of 6 different members, are known to regulate numerous pathways in different stem cell types [21]. GPCs present a core protein to which glycosaminoglycan chains are covalently linked. They are anchored to the cell membrane by a glycosylphosphatidylinositol (GPI) anchor [22,23]. The cleavage and/or the sulfation of GPCs play pivotal roles in their mechanism to regulate growth factor sequestration [24-27]. GPCs trigger a stimulatory [28] or inhibitory effect [29] according to the sequestrated growth factors.
Despite these observations, the link between HSPGs expressed by HF stem cells and physiological regulation of the hair growth cycle and hair shaft regeneration has not yet been established. However, some studies have highlighted the link between the alteration of the proteoglycan expression and/or the distribution and hair growth disorder such as androgenic alopecia (possibly due to the fact that androgens are able to modify the HSPG expression) [21]. Moreover, few studies have shown the beneficial effects of proteoglycan-based treatments against hair loss [30-34] and there are evidence that oral administration or application of proteoglycans can regulate some signaling pathways and modulate the cell fate [21]. The mechanism of action of HSPGs on signaling pathway involved in hair follicle stem cell fate remains to be elucidated in order to develop HSPGbased treatment for hair loss.
The aim of the present study was to identify the GPCs involved in the differentiation of HFSC and their role in the expression of BMP, Wnt and Shh and their receptors. A better understanding of these mechanisms could lead to the development of treatment for androgenic alopecia by targeting HSPGs.
Materials and Methods
Cell Culture
The cell types are described in Table 1. All cells were cultured at 37°C with 5% CO2 and used from passages 1 to 4 throughout the study.
Spheroid Preparation
The preparation of spheroids was performed by seeding the human hair follicle dermal papilla cells (HHFDPCs) in ultra-low attachment plates (40 000 cells/well). After 24 h, spheroids were formed, and they were maintained for several days in the same medium.
Conditioned Media Collection
After reaching 70% of confluence, HHFDPCs, keratinocytes of ORS (KORS) and human dermal microvascular endothelial cells (HDMECs) were starved by incubation in the hair follicle stem cell medium (HFSCM) without FBS and growth factors. The conditioned media were collected after 24 or 48 h and stored at -80°C for further analyses.
Differentiation Assay
Hair follicle stem cells (HFSCs) were seeded on 6- well plates (0.25 x 105 cells/mL) and incubated for 24 h in HFSCM. The medium of interest was then added and incubated for 5 days. After this time, the HFSCs were collected for further experiments.
Heparan Sulfate GAG Chain Cleavage by Heparinase III Treatment
First, the HFSCs were pre-incubated in the basal HFSC medium or in the medium of interest supplemented with 0.05 unit per mL (U/mL) of heparinase III, for 1 h at 37°C. Then, the pre-incubation medium was removed and medium supplemented with 0.01 U/mL of heparinase III was added till the end of the experiments.
Real-Time Reverse Transcription-Polymerase Chain Reaction
Using the PureLinkTM RNA mini kit (Thermo Fisher Scientific, Waltham, MA, USA), total RNA was extracted. Then, using the Maxima first-strand cDNA synthesis kit with dsDNase (Thermo Fisher Scientific, Waltham, MA, USA), 250 ng of total RNA were reverse-transcribed into cDNA. The Maxima SyBr green/ROX kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to perform real-time q-PCR. The primers used in this study are presented in Table 2. The fluorescence detection was carried out using Agilent MX300P device and MxPro software (Santa Clara, CA, USA). The ΔCt method was used to calculate the relative gene expression. According to the method of Kozera and Rapacz [35], the Ct (threshold cycle) of the gene of interest was compared with average of the Ct of two different reference genes: glyceraldehyde-3-phosphate dehydrogenase A (GAPDH) and TATA binding protein (TBP). Relative quantitative expression was determined as 2–ΔCt.
Immunobloting
Cell lysates were obtained using RIPA Buffer (Sigma-Aldrich, Saint-Louis, MO, USA) supplemented with 1% protease inhibitor cocktail. After 20 min of incubation on ice with regular mix every 5 min, cell lysates were precipitated by centrifugation at 10 000 g for 10 min at 4°C, and the supernatant of protein extract was collected.
Then, the samples were loaded in polyacrylamide gels as previously described [36]. The primary antibodies used are presented in Table 3.
Co-immunoprecipitation
For co-immunoprecipitation assays, Sepharose beads (protein A-Sepharose® 4B, Sigma-Aldrich) were used as previously described [37]. The primary antibodies used for these experiments are presented in Table 3.
Statistical Analysis
Results are normalized to the control condition and expressed as mean +/- SD. Statistical analyses were performed using SigmaPlot. Experiments were analyzed using Mann-Whitney U test. A p value less than 0.05 was considered significant. The respective p values are indicated in the figures as follows: *p < 0.05, **p < 0.01, ***p < 0.001.
Results
Role of KORS and HDMEC in the Regulation of the Balance Between the Wnt and BMP Expression in the HHFDPC Spheroids
The expression of Wnt and BMP was analyzed on HHFDPCs incubated or not with KORSCM or HDMECCM (Figure 1).
The gene expression of BMP2, 4 and 6 and Wnt3a and 5a in HHFDPCs was modulated by their incubation in the CM compared to HFSCM (Figure 1a). Particularly, the HDMECCM decreased the BMP2, 4 and 6 gene expression and increased the Wnt3a and 5a gene expression in HHFDPCs compared to the control condition in HFSCM. In contrast, the KORSCM only decreased the BMP2 gene expression and did not increase the Wnt3a and 5a gene expression in HHFDPCs compared to the control condition in HFSCM. Then, the protein expression of BMP2, 4 and 6 and Wnt3a and Wnt5a was analyzed in HHFDPCs (Figure 1b). The BMP2, 4 and 6 protein expression was strongly increased in HHFDPCs cultured in KORSCM. In contrast, when the HHFDPCs were incubated in the HDMECCM, BMP4 and 6 protein expression was decreased. The Wnt3a gene and protein expression was increased in HHFDPCs incubated in HDMECCM, compared to the HHFDPCs cultured in the HFSCM. The Wnt3a protein expression was increased in HHFDPCs incubated in KORSCM but not at the transcriptional level. The Wnt5a gene and protein expression was only increased when the HHFDPCs were incubated in the HDMECCM. These results are summarized in Figure 1c.
Effect of the Conditioned Media of HHFDPC Spheroids, KORS and HDMECs on HFSC Differentiation
Then, the direct effect of HHFDPC spheroids, KORS and HDMECs on HFSC differentiation was investigated. The expression of different markers of HF stem cells and progenitors was analyzed (Figure 2). According to the specific expression pattern of these markers described in the literature (Figure 2a), the effects of HHFDPC spheroids-conditioned medium (HHFDPCCM), KORSCM and HDMECCM on HFSC differentiation were investigated. The Figure 2b presents the results of the analyses of the gene expression of the HFSC markers. When cultured in specific HFSC medium (HFSCM), HFSCs presented a high expression of ITGB1, ITGA6, cK15, CD200 and CD34 but not LGR5. Interestingly, when HFSC were incubated in the three different CM, the expression of ITGA6, cK15 and CD200 decreased. A strong increase of LGR5 gene expression was observed under HHFDPCCM condition. In contrast, the KORSCM led to an increase of CD34 gene expression and a decrease of ITGB1 gene expression. For protein expression, analyses were focused on CD34 and CD200/LGR5 expression because they are specific biomarkers of ORS progenitor cells and SHG progenitor cells, respectively (Figure 2a). The analysis of CD200/LGR5 protein expression showed that only the HHFDPCCM increased the expression of these both markers in HFSCs compared to the control medium HFSCM (Figure 2c). Interestingly, the gene expression of CD200 was decreased when HFSC were incubated in HHFDPCCM. In contrast, only the KORSCM induced a strong increase of the expression of CD34 and the HDMECCM to moderately increase CD34 expression in HFSCs, both compared to the HFSCM control medium. Moreover, when HFSCs were incubated in HDMECCM, a decreased expression of CD200 was observed compared to the control HFSCM condition.
Involvement of BMP, Wnt, and Shh in Intercellular Communication
The protein expression of BMP2, 4 and 6, Wnt3a and 5a, and Shh in KORS and HDMECs was investigated (Figure 3). BMP2, 4 and 6 were detected in KORS, while only BMP6 was detected in HDMECs (Figure 3a). Wnt3a and 5a were not detected in KORS and HDMECs (data not shown). These two cell types were demonstrated to express Shh (Figure 3b). Indeed, the expression of the monovalent Shh (49 and 51 kDa) as well as a multimeric form at 140 kDa was detected in KORS. In contrast, a unique multimeric form at 70 kDa was observed in HDMECs. In comparison, Shh was not detected in HHFDPCs (data not shown). However, these cells presented a strong gene expression of Wnt5a (in contrast to Wnt3a), a moderate gene expression of BMP2, and a low gene expression of BMP4 and 6 (Figure 3c). The protein expression of BMPs and Wnts by the HHFDPC spheroids has been presented in Figure 1b. The Figure 3d shows the balance of the results obtained by the gene and protein analyses.
Role of GPC1, 4 and 6 as Co-receptors of BMPR2,Fz7 and Patched1 and Potential Involvement in the Differentiation of HFSCs
The interactions of GPC1, 4 and 6 with the receptors of BMP (BMPR2), Wnt (Fz7), and Shh (Patched1) were investigated in HFSCs by co-immunoprecipitation (Figure4).
After GPC1, 4 or 6 immunoprecipitations, BMPR2, Fz7, and Patched1 were detected by immunoblotting with different protein level (Figure 4a-c). BMPR2 displayed a slightly higher interaction level with GPC4 and 6 compared to GPC1 (Figure 4a). GPC1 and 4 strongly interact with Fz7 (Figure 4b) and Patched1 (Figure 4c) compared to GPC6. Moreover, conversely, after immunoprecipitation with BMPR2, Fz7 or Patched1 antibody, GPC1, 4 and 6 were detected by immunoblotting (Figure 4d-f). GPC1 and 4 immunoprecipitated preferentially with Fz7 in comparison with BMP and Patched1 while GPC6 exhibited similar affinity with the three receptors.
Impact of Heparinase III on HFSC Differentiation Induced by HHFDPC Spheroids-, KORS-, and HDMEC-Conditioned Media
To provide another evidence of the involvement of GPCs in the regulation of HFSC differentiation induced by growth factors secreted by HHFDPC, KORS and/or HDMEC, the effect of HS glycosaminoglycan chain cleavage by heparinase III was assessed on HFSC differentiation (Figure5).
The heparinase III treatment increased the protein expression of LGR5 in HFSCs and with HHFDPCCM, in particular. In contrast, the heparinase III treatment induced a decrease of CD34 expression in HFSCs incubated in the three CM, and more specifically in KORSCM and HDMECCM which were shown to induce
Discussion
The study was devoted to characterizing the cell communications of KORS, HHFDPC and HDMECs with HFSCs for a better understanding of the regulation of HFSC fate. Indeed, HFSC behavior, in vivo, is known to be regulated by the dermal papilla through BMP and Wnt secretion and by the hair matrix through Shh secretion [11,12]. To our knowledge, the potential role of KORS and endothelial hair cells in the regulation of HFSC differentiation remains unclear. Here, the role of these two cell types was investigated in order to determine their effect on (i) indirect HFSC differentiation by modulating the regulation of the balance between the Wnt and BMP expression in the HHFDPC spheroids and on (ii) direct HFSC differentiation.
The dermal papilla is known to secrete Wnt and BMP in order to regulate the HF stem cell behavior during the hair growth cycle [11,12]. The temporal and spatial course of the dermal papilla activation leading to the telogen-anagen transition is not fully understood yet. It is known that macrophages and adipocytes have a role in the telogen-anagen transition associated with the activation of Wnt secretion by the dermal papilla [38,39] but the potential role of KORS or endothelial cells is still not described to our knowledge.
It is known that the dermal papilla secretes Wnts,once activated by adipocytes at the beginning of anagen phase, and BMPs during catagen / telogen phases [12]. All the light has not yet been shed on the activation cycle of dermal papilla and the balance between Wnts or BMPs expression during the different phases of hair growth cycle.
In the present study, the results suggest the involvement of KORS and HDMEC in the regulation of the growth factors expressed by HHFDPC. In particular, HHFDPCs presented an increased expression of Wnt3a and 5a after incubation in HDMECCM. Moreover, the expression of BMP4 and 6 decreased in HHFDPCs after incubation in HDMECCM. These results highlighted the probable role of endothelial cells in the activation of dermal papilla cells during telogen to anagen transition. Interestingly, the incubation in KORSCM led to an increase of BMP2, 4 and 6 protein expression pointing out the probable role of the keratinocytes of the ORS in the resumption of BMP expression by the dermal papilla during anagen to catagen transition. Although, BMP2 protein increased independently of the mRNA expression, suggesting a post-translational regulation.
The stimulatory effect of HDMECCM on the expression of Wnt3a and Wnt5a was observed with a concomitant decreased expression of BMP4 and 6 in HHFDPC. Interestingly enough, this induction of Wnts observed concomitantly with a decreased of BMP expression was already described during telogen to anagen transition in the literature [11,12]. Altogether, this observation suggests a key of HDMEC on the telogen to anagen transition. Our results obtained with KORSCM also suggest that KORS might take over by stimulating the BMP2, 4 and 6 expression by the dermal papilla during anagen to catagen transition. Thus, the HHFDPC could represent an interesting source of specific differentiation factors which composition might specifically differs according to the interaction of HHFDPC with HDMEC or KORS.
Wnt and BMP are known to be secreted by the dermal papilla and to regulate HFSC behavior. Thus, the effect of Wnt and BMP expression in HHFDPC spheroid by HDMECCM and KORSCM highlights the probable indirect role of these two cell types on the regulation of HFSC differentiation.
Then, the direct effect of the conditioned media of HHFDPC spheroids, KORS and HDMECs on HFSC differentiation was investigated, according to the specific expression pattern of their markers described in the literature.
The results obtained, demonstrated that the HFSC line used for this study presented the same characteristics as the human hair follicle stem cells described in the literature. Moreover, the results underlined the paracrine communication of KORS, HHFDPCs and HDMECs with HFSCs. Indeed, by following the expression of CD34 and LGR5/CD200 specific biomarkers, the three CM were shown to induce different level of differentiation of HFSC. In particular, HHFDPCCM seems to promote the HFSC differentiation towards a SHG progenitor phenotype while the KORSCM appears to lead to the differentiation of HFSCs towards an ORS progenitor phenotype. In literature, it is known that the dermal papilla plays an important role in the induction of hair follicle differentiation [1,40]. Our results are in accordance with this point. Moreover, they provide new insights on the potential role of KORS on this process. The HDMECCM seemed to favor the appearance of intermediate phenotypes of HFSC differentiation. Indeed, when cultured in HDMECCM, HFSCs expressed all markers, including CD34, compared to HFSCs incubated in HFSCM or HHFDPCCM which are CD34 negative. Moreover, in HDMECCM, HFSCs presented a moderate level of CD34 expression, compared to HFSCs in KORSCM, and a moderate level of CD200/LGR5 expression, compared to HFSCs in HHFDPCCM. Interestingly, the gene expression of CD200 was decreased, contrary to the protein expression, when HFSC were incubated in HHFDPCCM, suggesting a post-translational mechanism. Numerous phenotypes reflecting the different level of hair follicle stem cells are described in the literature. In a simplified way, five large distinct populations are defined in the literature (see introduction) but intermediate phenotypes are also reported [5-9]. These results suggest that endothelial cells may play a role in the early stage of HFSC differentiation.
Three major growth factors are described in the literature to regulate HFSC differentiation: Wnts, BMPs and Shh [11-13]. The expression of these growth factors was analyzed in KORS, HDMECs and HHFDPCs to correlate or not the effect of the three CM on HFSC differentiation with these growth factors.
The present report demonstrates that KORS, HDMECs and HHFDPCs express at least one growth factor promoting stem cell proliferation/differentiation (Wtn3a, Wnt5a or Shh) and at least one growth factor promoting quiescence (BMP2, 4 or 6), explaining, at least in part, their effects on stem cells differentiation (see Figure 3d). The fact that KORS and HDMECs do not express Wnt3a or Wnt5a (not shown) might explain why CM of these cells do not induce the differentiation of bulb stem cells into SHG progenitors (CD200/LGR5 markers). Indeed, Wnts are known to induce the differentiation of bulb stem cells into SHG progenitors [11,12]. On the other hand, only KORS and HDMECs secrete Shh (in different forms) and their CM are the only ones to promote the differentiation of bulge stem cells into ORS progenitors (CD34 marker). In the literature, Shh is described to be secreted by matrix cells and to stimulate the differentiation of bulge stem cells to provide a new pool of ORS progenitor cells [13]. Taking into account the localization of bulb stem cells in the ORS at the early anagen phase, it is not surprising that KORS may also be involved in the induction of bulb stem cell differentiation into ORS progenitor cells through Shh secretion. On the contrary, HHFDPCCM which has been shown to induce bulge stem cell differentiation into SHG progenitors, expressed Wnt3a and Wnt5a but not Shh.
HSPGs such as GPCs play an important role in the regulation of stem cell behavior by modulating signaling pathways associated with Wnt, BMP or Shh [33,34]. The interaction of GPCs with the receptors associated with these three growth factors was investigated in the differentiation of HFSCs. Indeed, GPCs are GPI-anchored membrane HSPGs known to interact with their core protein or their GAG chains with several receptors and/or growth factors to regulate the associated signaling pathways [20,22,27]. Based on this knowledge, the interactions of GPC1, 4 and 6 with the receptors of BMP (BMPR2), Wnt (Fz7), and Shh (Patched1) were investigated in HFSCs by co-immunoprecipitation assays (Figure 4).
The results suggest that GPC1, 4 and 6 may regulate the balance between quiescence and differentiation of HFSCs by the formation of tripartite complexes with Wnt and their Fz receptors, with BMPs and their BMP receptors as well as with Shh and their Patched receptors. The association between these receptors and GPC1, 4 and 6 has been already probed in numerous other tissues and organs where the role of GPCs in the regulation of stem cell behavior has been demonstrated [29,41-45].
The fact that an association of GPC1, 4 and 6 with the receptors of the growth factors involved in HFSC behavior (BMP, Wnt and Shh) occurs in bulge stem cells and highlights the probable role of these three GPCs in the regulation of HFSC behavior modulated by BMP, Wnt and Shh.
GPCs exhibit HS chains known to be involved in the interaction of GPCs with receptor and/or growth factors to regulate the associated signaling pathways [29,30,46]. To provide another evidence of the involvement of GPCs in the regulation of HFSC differentiation induced by growth factors secreted by HHFDPC, KORS and/or HDMEC, the effect of HS glycosaminoglycan chain cleavage by heparinase III was assessed on HFSC differentiation.
The results provided evidence on the role of HS chains in the regulation of HFSC fate. Indeed, in the absence of HS chains, hair follicle stem cells appear to differentiate towards an exclusively secondary germ progenitor profile and provide fewer ORS progenitors. These results may explain the loss of proliferation/differentiation potential of hair follicle stem cells observed in androgenetic alopecia [47,48]. The growth factors secreted by the dermal papilla, the keratinocytes of the ORS and/or the endothelial cells of the HF, seem to no longer act in a controlled manner on the hair follicle stem cells. Their differentiation is disturbed, and the progenitor cells of the ORS are less produced. Targeting GPCs to improve their sulfation could lead to a restauration of the action of Wnt, BMP and/or Shh pathway and the formation of progenitor cells of the ORS and, thus, a potential treatment for androgenic alopecia.
All these results demonstrate the involvement of GPCs in the regulation of hair stem cell differentiation and highlight the complexity of the mechanism of regulation of the stem cell fate. Indeed, some studies have shown the importance of HS chains in the regulation of some growth factors [24-27]. Here, the involvement of HS is strongly suggested regarding the results of the cleavage of HS chains. e study of the stem cell fate regulation through the modulation of signal pathways by GPCs is complex and has to take into account that GPCs act differently according to their sulfation and/or their cleavage. However, the study of these mechanisms is crucial to develop treatments targeting stem cells, such as in dermatological studies (androgenic alopecia) or cancerological studies.
Conclusion
This report provides more information on the intercellular paracrine communication of HHFDPC, KORS or HDMEC with HFSCs. Indeed, the results suggest the involvement of both KORS and HDMEC in the regulation of HFSC differentiation through the secretion of BMPs and Shh. Moreover, the analysis of the expression of BMPs and Wnts in HHFDPC underlines the potential role of HDMEC in the induction of Wnt expression in HHFDPC and the potential role of KORS in the induction of BMP expression in HHFDPC. Finally, the association of GPC1, 4 and 6 with BMPR2, Fz7 and Patched 1 as well as the effect of HS chain cleavage on HFSC differentiation highlight the probable role of GPC1, 4 and 6 as coreceptors regulating the hair stem cell fate.
Acknowledgments
This study was made in collaboration with BASF Beauty Care Solutions.
- G Cotsarelis, TT Sun, RM Lavker (1990) Label-retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis, Cell, 61: 1329-37.
- A Sada, T Tumbar (2013) New insights into mechanisms of stem cell daughter fate determination in regenerative tissues, Int Rev Cell Mol Biol. 300: 1-50.
- P Myung, M Ito, (2012) Dissecting the bulge in hair regeneration, J Clin Invest. 122: 448-54.
- L Alonso, E Fuchs (2003) Stem cells of the skin epithelium, Proceedings of the National Academy of Sciences, 100: 11830-5.
- AA Panteleyev (2018) Functional anatomy of the hair follicle: The Secondary Hair Germ, Experimental Dermatology, 27: 701-20.
- M Ohyama, A Terunuma, CL Tock, MF Radonovich, CA Pise-Masison, SB Hopping, et al. (2006) Characterization and isolation of stem cell–enriched human hair follicle bulge cells, J Clin Invest. 116: 249-60.
- JE Kloepper, S Tiede, J Brinckmann, DP Reinhardt, W Meyer, R Faessler, R Paus (2008) Immunophenotyping of the human bulge region: the quest to define useful in situ markers for human epithelial hair follicle stem cells and their niche, Experimental Dermatology, 17: 592-609.
- K Inoue, N Aoi, T Sato, Y Yamauchi, H Suga, H Eto, H Kato, et al. (2009) Yoshimura, Differential expression of stem-cell-associated markers in human hair follicle epithelial cells, Lab. Invest. 89: 844-56.
- TS Purba, IS Haslam, E Poblet, F Jiménez, A Gandarillas, A Izeta, R Paus (2014) Human epithelial hair follicle stem cells and their progeny: current state of knowledge, the widening gap in translational research and future challenges, Bioessays, 36: 513-25.
- VA Botchkarev, R Paus (2003) Molecular biology of hair morphogenesis: development and cycling, J. Exp. Zool. B Mol. Dev. Evol. 298: 164-80.
- MV Plikus, RE Baker, CC Chen, C Fare, D de la Cruz, et al. (2011) Self-organizing and stochastic behaviors during the regeneration of hair stem cells, Science, 332: 586-9.
- R Sennett, M Rendl (2012) Mesenchymal-epithelial interactions during hair follicle morphogenesis and cycling, Semin. Cell Dev. Biol. 23: 917-27.
- YC Hsu, L Li, E Fuchs (2014) Transit-amplifying cells orchestrate stem cell activity and tissue regeneration, Cell, 157: 935-49.
- BA Bernard (2016) Advances in Understanding Hair Growth, F1000Res 5 (2016) F1000 Faculty Rev-147.
- VA Botchkarev, J Kishimoto (2003) Molecular control of epithelial-mesenchymal interactions during hair follicle cycling, J Investig Dermatol Symp Proc. 8: 46-55.
- JR Couchman (1993) Hair follicle proteoglycans, J Invest Dermatol. 101: 60S-4.
- S Malgouries, S Thibaut, BA Bernard (2008) Proteoglycan expression patterns in human hair follicle, Br. J. Dermatol. 158: 234-42.
- GE Westgate, AG Messenger, LP Watson, WT Gibson (1991) Distribution of proteoglycans during the hair growth cycle in human skin, J Invest Dermatol, 96: 191-5.
- IB Bayer-Garner, RD Sanderson, BR Smoller (2002) Syndecan-1 is strongly expressed in the anagen hair follicle outer root sheath and in the dermal papilla but expression diminishes with involution of the hair follicle, Am J Dermatopathol. 24: 484-9.
- Karamanos NK, Piperigkou Z, eocharis AD, Watanabe H, Franchi M, et al. (2018) Proteoglycan Chemical Diversity Drives Multifunctional Cell Regulation and erapeutics. Chemical Reviews, 118: 9152-232.
- Colin-Pierre C, El Baraka O, Danoux L, Bardey V, André V, Ramont L, Brézillon S (2022) Regulation of stem cell fate by HSPGs: implication in hair follicle cycling. NPJ Regenerative medicine, 7: 77.
- Filmus J, Capurro M, Rast J (2008) Glypicans.Genome Biology, 9: 224
- U Häcker, K Nybakken, N Perrimon (2005) Heparan sulphate proteoglycans: the sweet side of development, Nat Rev Mol Cell Biol. 6: 530-41.
- KL Ayers, A Gallet, L Staccini-Lavenant, PP Thérond (2010) The long-range activity of Hedgehog is regulated in the apical extracellular space by the glypican Dally and the hydrolase Notum, Dev. Cell, 18: 605-20.
- J Kreuger, L Perez, AJ Giraldez, SM Cohen (2004) Opposing activities of Dally-like glypican at high and low levels of Wingless morphogen activity, Dev. Cell, 7: 503-12.
- DA Pye, RR Vives, JE Turnbull, P Hyde, JT Gallagher (1998) Heparan sulfate oligosaccharides require 6-O-sulfation for promotion of basic fibroblast growth factor mitogenic activity, J. Biol. Chem. 273: 22936-42.
- J Filmus (2023) Glypicans, 35 years later, Proteoglycan Research, 1: e5.
- S Yamamoto, H Nakase, M Matsuura, Y Honzawa, K Matsumura, N Uza, et al. (2013) Heparan sulfate on intestinal epithelial cells plays a critical role in intestinal crypt homeostasis via Wnt/β-catenin signaling, Am. J. Physiol. Gastrointest. Liver Physiol. 305: G241-9.
- PP Dwivedi, RH Grose, J Filmus, CST Hii, CJ Xian, PJ Anderson, BC Powell, (2013) Regulation of bone morphogenetic protein signalling and cranial osteogenesis by Gpc1 and Gpc3, Bone, 55: 367-76.
- J Wadstein, E Thom, A Gadzhigoroeva (2020) Integral Roles of Specific Proteoglycans in Hair Growth and Hair Loss: Mechanisms behind the Bioactivity of Proteoglycan Replacement Therapy with Nourkrin® with Marilex® in Pattern Hair Loss and Telogen Effuvium, Dermatol Res Pract.
- A Lassus, L Jeskanen, HP Happonen, J Santalahti (1991) Imedeen for the treatment of degenerated skin in females, J Int Med Res. 19: 147-52.
- E Thom (2001) Efficacy and tolerability of Hairgain in individuals with hair loss: a placebo-controlled, double-blind study, J Int Med Res. 29: 2-6.
- E Thom (2006) Nourkrin: objective and subjective effects and tolerability in persons with hair loss, J Int Med Res. 34: 514-9.
- DH Kingsley, E Thom (2012) Cosmetic hair treatments improve quality of life in women with female pattern hair loss, J Appl Cosmetol. 30: 49-59.
- Kozera B, Rapacz M (2013) Reference genes in realtime PCR. Journal of Applied Genetics, 54: 391-406.
- G Perrot, C Colin-Pierre, L Ramont, I Proult, C Garbar, V Bardey, et al. (2019) Decreased expression of GPC1 in human skin keratinocytes and epidermis during ageing, Exp. Gerontol. 126: 110693.
- C Colin-Pierre, N Berthélémy, N Belloy, L Danoux, V Bardey, R Rivet, et al. (2021) The Glypican-1/HGF/C-Met and Glypican-1/VEGF/VEGFR2 Ternary Complexes Regulate Hair Follicle Angiogenesis, Front Cell Dev Biol. 9: 781172.
- ECE Wang, CA Higgins (2020) Immune cell regulation of the hair cycle, Experimental Dermatology, 29: 322-33.
- E Festa, J Fretz, R Berry, B Schmidt, M Rodeheffer, M Horowitz, V Horsley (2011) Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling, Cell, 146: 761-71.
- HB Chase (1954) Growth of the hair, Physiol. Rev. 34: 113-26.
- M Capurro, T Izumikawa, P Suarez, W Shi, M Cydzik, T Kaneiwa, et al. (2017) Glypican-6 promotes the growth of developing long bones by stimulating Hedgehog signaling, J. Cell Biol. 216: 2911-26.
- J Filmus, M Capurro (2014) The role of glypicans in Hedgehog signaling, Matrix Biol. 35: 248-52.
- A Fico, A De Chevigny, J Egea, MR Bösl, H Cremer, F Maina, R Dono (2012) Modulating Glypican4 suppresses tumorigenicity of embryonic stem cells while preserving self-renewal and pluripotency, Stem Cells. 30: 1863-74.
- M Venero Galanternik, ME Lush, T Piotrowski (2016) Glypican4 modulates lateral line collective cell migration non cell-autonomously, Dev Biol. 419: 321-35.
- W Shi, T Kaneiwa, M Cydzik, J Gariepy, J Filmus (2020) Glypican-6 stimulates intestinal elongation by simultaneously regulating Hedgehog and non-canonical Wnt signaling, Matrix Biol. 88: 19-32.
- P Jakobs, P Schulz, C Ortmann, S Schürmann, S Exner, R Rebollido-Rios, et al. (2016) Bridging the gap: heparan sulfate and Scube2 assemble Sonic hedgehog release complexes at the surface of producing cells, Sci Rep. 6: 26435.
- LA Garza, CC Yang, T Zhao, HB Blatt, M Lee, H He, et al. (2011) Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200-rich and CD34-- positive hair follicle progenitor cells, J. Clin. Invest. 121: 613-22.
- Y Mao, P Liu, J Wei, Y Xie, Q Zheng, R Li, J Yao (2023) Cell Therapy for Androgenetic Alopecia: Elixir or Trick?, Stem Cell Rev Rep. 19: 1785-99.
FIGURE 1
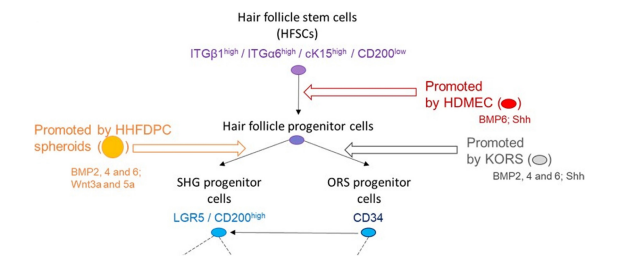
FIGURE 2
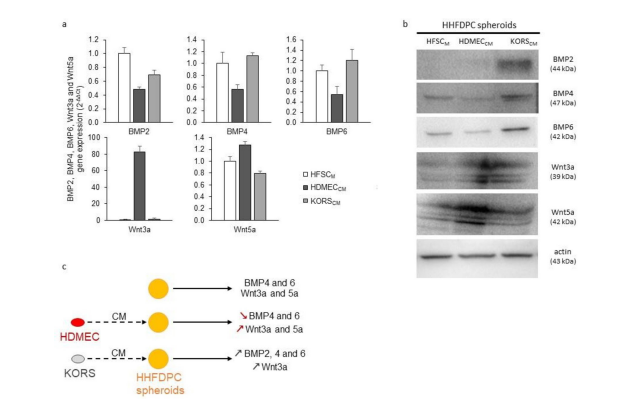
Figure 1: Effect of KORSCM and HDMECCM on growth factor expression by HHFDPCs. (a-b) Gene (a) and protein (b) expressions of BMP2, BMP4, BMP6, Wnt3a and Wnt5a in HHFDPCs.These cells were incubated in KORSCM or HDMECCM for 48 h before the analysis. N=1 experiment, n=3 replicates. (c) A summary diagram is presented to illustrate the effect of the conditioned media on the expression of BMP2, 4 and 6 as well as Wnt3a and 5a by HHFDPCs.
FIGURE 3
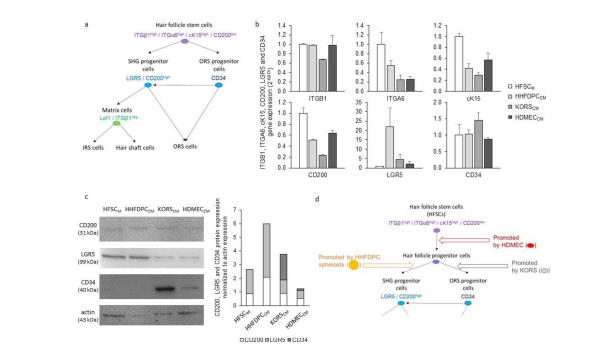
Figure 2: Effect of HHFDPCCM, KORSCM and HDMECCM on HFSC diffrentiation (a) A scheme of HFSC diffrentiation, as described in the literature, is illustrated. (b) ITGB1, ITGA6, cK15, CD200, LGR5 and CD34 gene expression in HFSCs was analyzed by RT-qPCR. N=2 independent experiments, n=3 replicates per experiment. (c) Protein expression and quantification of CD200, LGR5 and CD34 in the HFSCs was determined by Western immunoblotting.These HFSCs were incubated in HHFDPCCM, KORSCM or HDMECCM for 48 h before the analysis. (d) A summary diagram is presented to illustrate the effect of the conditioned media on HFSCs diffrentiation.
FIGURE 4
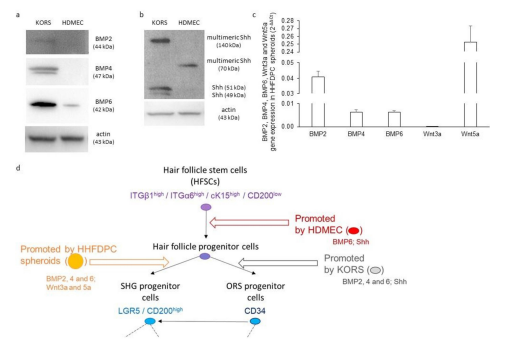
Figure 3: KORS, HDMECs and HHFDPCs express growth factors known to be involved in the regulation of HFSC behavior. (a-b) Protein expression of BMP2, BMP4, BMP6 (a) and Shh (b) in the HFSCs was determined by Western immunoblotting. (c) BMP2, BMP4, BMP6, Wnt3a and Wnt5a gene expression in HHFDPC spheroids were analyzed by RT-qPCR. N=2 independent experiments, n=3 replicates per experiment. (d) A summary diagram is presented to illustrate the effect of the conditioned media on HFSCs, and the growth factors expressed by KORS, HDMECs and HHFDPCs.
FIGURE 5
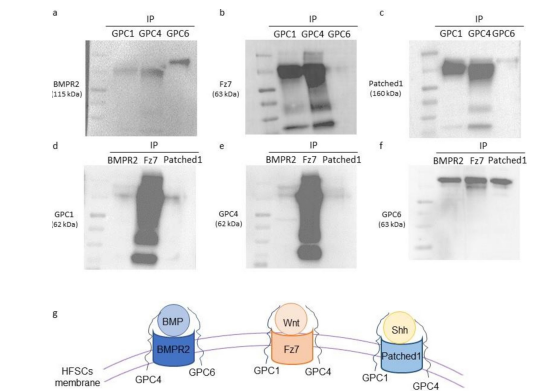
Figure 4: GPC1, GPC4 and GPC6 interact with BMPR2, Fz7 and Patched 1. (a-c) A whole cell protein extract (25 μg) from the HFSCs was precipitated using anti-GPC1, anti-GPC4 or anti-GPC6 antibody. Anti-BMPR2 (a), anti-Fz7 (b) or anti-Patched 1 (c) antibodies were used to reveal the membrane. (d-f) A reverse immunoprecipitation assay, corresponding to the co-immunoprecipitation shown in Figure 4a-c, was conducted from HFSC whole cell protein extracts using anti-BMPR2, anti-Fz7 or anti-- Patched 1 antibody. Then, the isolated immunocomplexes were immunoblotted using anti-GPC1 (d), anti-GPC4 (e) or anti-GPC6 (f) antibodies. (g) A summary diagram is presented to illustrate the interaction of GPC1, 4 and 6 with BMPR2, Fz7 and Patched 1.
FIGURE 6
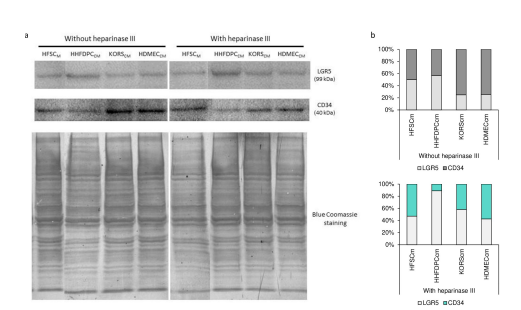
Figure 5: Effect of heparinase III incubation on HFSC diffrentiation induced by HHDCFCM, KORSCM or HDMECCM. Protein expression and quantification of LGR5 and CD34 in the HFSCs was determined by Western immunoblotting.The cells were incubated in HHFDPCCM, KORSCM or HDMECCM without or with heparinase III treatment for 48 h before the analysis.
Tables at a glance
Figures at a glance